Функциональное значение базальных ядер головного мозга. Функции базальных ядер. Строение и расположение
Одной из самых необъяснимых вещей во Вселенной является мозг. О нём почти ничего неизвестно, что касается принципов функционирования. С точки зрения физиологии, этот орган неплохо изучен, однако большинство людей имеют более чем поверхностное представление о его строении.
Преимущественному числу образованных людей известно, что головной мозг – это два полушария, покрытые корой да извилинами, условно состоит он из нескольких отделов и где-то там есть серое и белое вещества. Обо всём этом мы расскажем в специальных темах, а сегодня рассмотрим, что такое базальные ядра головного мозга, о которых слышали и знают немногие.
Строение и расположение
Базальные ганглии головного мозга – скопление серого вещества в белом, расположенное у основания головного мозга и входящее в состав его передней доли. Как видим, серое вещество не только полушария формирует, но и находится в виде отдельных скоплений, называемых ганглиями. Они имеют тесную связь с белым веществом и корой обоих полушарий.
Строение этой области основано на срезе головного мозга. В её состав входят:
- миндалевидное тело;
- полосатое тело (в состав входят хвостатое ядро, бледный шар, скорлупа);
- ограда;
- чечевицеобразное ядро.

Между чечевицеобразным ядром и таламусом находится белое вещество, называемое внутренней капсулой, между островком и оградой – наружная капсула. В последнее же время предлагается немного иная структура подкорковых ядер головного мозга:
- полосатое тело;
- несколько ядер среднего и промежуточного мозга (субталамическое, ножкомостовое и чёрная субстанция).
Вместе они отвечают за двигательную активность, координацию движений и мотивацию в человеческом поведении. Это всё, что можно наверняка сказать о функции подкорковых ядер. В остальном они, как и мозг в целом, малоизучены. О назначении оградки известно ровно ничего.
Физиология
Все подкорковые ядра опять же условно объединяются в две системы. Первая называется стриопаллидной системой, в состав которой включены:
- бледный шар;
- хвостатое ядро головного мозга;
- скорлупа.
Две последние структуры состоят из множества слоёв, благодаря чему их сгруппировали под именем стриатум. Бледный шар отличается более ярким, светлым цветом и не является слоистым.
Чечевицеобразное ядро образуется бледным шаром (располагается внутри) и скорлупой, которая образовывает её наружный слой. Ограда с миндалевидным телом являются составляющими лимбической системы головного мозга.
Рассмотрим подробнее, что собой представляют эти ядра мозга.
Хвостатое ядро
Парная составляющая головного мозга, относящаяся к полосатому телу. Место локализации – впереди от таламуса. Их отделяет полоса белого вещества, называемая внутренней капсулой. Его передняя часть имеет более массивную утолщённую структуру, головка структуры примыкает к чечевицеобразному ядру.
По структуре оно состоит из нейронов Гольджи и имеет следующие характеристики:
- их аксон очень тонкий, а дендриты (отростки) – короткие;
- нервные клетки имеют уменьшенные, в сравнении с нормальными, физические размеры.
Хвостатое ядро имеет тесные связи с множеством иных выделенных структур мозга и образует очень широкую сеть нейронов. Через них бледный шар и таламус взаимодействуют с сенсорными участками, создавая пути с замкнутыми контурами. Взаимодействует ганглий и с иными участками мозга, причём не все они лежат по соседству с ним.

Специалисты не имеют общего мнения по поводу того, какова функция хвостатого ядра. Это ещё раз подтверждает необоснованную, с научной точки зрения, теорию, что мозг является единой структурой, любая из его функций с лёгкостью выполняется любым участком. И это неоднократно доказано при исследованиях людей, пострадавших вследствие аварий, иных ЧП и болезней.
Наверняка известно, что он принимает участие в вегетативных функциях, играет важную роль в развитии познавательных способностей, координации и стимуляции двигательной активности.
Полосатое ядро состоит из чередующихся по большому счету в вертикальной плоскости слоев белого и серого веществ.
Чёрная субстанция
Составляющая системы, которая принимает наибольшее участие в координации движений и моторике, поддержке мышечного тонуса и управлении при соблюдении поз. Участвует во множестве вегетативных функций, таких как дыхание, сердечная деятельность, поддержка тонуса сосудов.
Физически субстанция является непрерывной полосой, как считалось на протяжении десятилетий, однако анатомические срезы показали, что она состоит из двух частей. Одна из них – приемник, который направляет дофамин полосатому телу, вторая – передатчик – служит транспортной артерией для передачи сигналов от базальных ганглиев иным отделам мозга, коих насчитывается более десятка.
Чечевицеобразное тело
Место его дислокации между хвостатым ядром и таламусом, кои, как говорилось, разделяются наружной капсулой. Спереди структуры она сливается с головкой хвостатого ядра, отчего её фронтальный срез имеет клинообразную форму.
Это ядро состоит из отделов, разделённых тончайшей плёнкой белого вещества:
- скорлупа – более темная наружная часть;
- бледный шар.
Последний сильно разнится со скорлупой строением и состоит из клеток Гольджи I-го типа, кои преобладают в человеческой нервной системе, и больше по размеру, чем их II-я разновидность. По предположениям нейрофизиологов, он является более архаичной мозговой структурой, чем иные составляющие ядра головного мозга.

Иные узлы
Ограда – тончайший слой серого вещества между скорлупой и островком, вокруг которого находится белая субстанция.
Также базальные ядра представляются и миндалевидным телом, находящимся под скорлупой в височной области головы. Считается, но наверняка не известно, что эта часть относится к обонятельной системе. В ней же заканчиваются нервные волокна, идущие из обонятельной доли.
Последствия нарушений физиологии
Отклонения в строении или функционировании ядер мозга сразу же приводят к следующим симптомам:
- движения становятся медленными и неуклюжими;
- нарушается их координация;
- появление произвольных сокращений и расслаблений мышц;
- тремор;
- невольное произношение слов;
- повторение однообразных несложных движений.

Фактически эти симптомы и дают понять о предназначении ядер, чего явно недостаточно, чтобы узнать об их истинных функциях. Периодически наблюдаются и проблемы с памятью. При наличии этих симптомов необходимо обратиться к врачу. Он назначит и процедур для проведения более точной диагностики в виде:
- ультразвукового исследования головного мозга;
- компьютерной томографии;
- сдачи анализов;
- прохождения специальных тестов.
Все эти меры помогут определить степень поражения, если оно есть, а также назначить курс лечения специальными препаратами. В некоторых ситуациях лечение может стать пожизненным.
К таким нарушениям относятся:
- дефицитарность ганглий (функциональная). Появляется у детей в связи с генетической несовместимостью их родителей (так называемое смешивание кровей разных рас и народов) и зачастую передаётся по наследству. В последнее десятилетие людей с подобными отклонениями все больше и больше. Возникает и у взрослых и перетекает в болезнь Паркинсона или Гентингтона, а также подкорковый паралич;
- киста базальных ганглий – результат неправильного обмена веществ, питания, атрофирования тканей мозга и воспалительных процессов в нём. Самым тяжелым симптомом является кровоизлияние в мозг, после которого вскоре наступает смерть. Опухоль хорошо различима на МРТ, не имеет тенденций к увеличению, не причиняет неудобств больному.
Базальные ядра, названные гистологами прошлого века ганглиями, представляют собой структуры ядерного типа, которые располагаются в толще белого вещества переднего мозга ближе к его основанию. У млекопитающих к базальным ядрам относятся сильно вытянутое в длину и изогнутое хвостатое ядро и заложенное в толще белого вещества чечевицеобразное ядро. Двумя белыми пластинками оно подразделяется на три части: наиболее крупную, лежащую латерально скорлупу, и бледный шар, состоящий из внутреннего и внешнего отделов (рис. 3.29).
Эти анатомические образования формируют так называемую стриопаллидарную систему (От лат. striatus - полосатый и pallidus - бледный.), которая по филогенетическим и функциональным критериям разделяется на древнюю часть палеостриатум и новую - неостриатум. Палеостриатум представлен бледным шаром, а неостриатум, появляющийся впервые у рептилий, состоит из хвостатого ядра и скорлупы, которые объединяются под названием полосатого тела, или стриатума. Хвостатое ядро и скорлупа связаны анатомически и характеризуются чередованием белого и серого вещества, что оправдывает возникновение термина полосатое тело.
К стриопаллидарной системе часто относят также субталамическое ядро (люисово тело) и черное вещество среднего мозга, которые образуют с базальными ядрами функциональное единство. Полосатое тело состоит главным образом из мелких клеток, аксоны которых направляются к бледному шару и черному веществу среднего мозга.
Полосатое тело является своеобразным коллектором афферентных входов, идущих к базальным ядрам. Главными источниками этих входов служат новая кора (преимущественно сенсомоторная), неспецифические ядра таламуса и дофаминергические пути от черного вещества.
В противоположность полосатому телу бледный шар состоит из крупных нейронов и является сосредоточением выходных, эфферентных путей стриопаллидарной системы. Аксоны локализованных в бледном шаре нейронов подходят к различным ядрам промежуточного и среднего мозга, в том числе и к красному ядру, где начинается красноядерно-спинномозговой путь экстрапирамидной системы двигательной регуляции.
Другой важный эфферентный путь идет от внутреннего отдела бледного шара к передневентральному и вентролатеральному ядрам таламуса, а оттуда продолжается к двигательным областям коры головного мозга. Наличие этого пути обусловливает многозвенную петлеобразную связь между сенсомоторными и двигательными областями коры, которая осуществляется через полосатое тело и бледный шар к таламусу. Примечательно, что в составе этого стриопаллидоталамокортикального пути базальные ядра выполняют роль афферентного звена по отношению к моторным областям коры головного мозга. Многочисленные связи стриопаллидарной системы с различными отделами мозга свидетельствуют об ее участии в процессах интеграции, однако до настоящего времени в знании о функциях базальных ядер остается много невыясненного.
Базальные ядра играют важную роль в регуляции движений и сенсомоторной координации. Известно, что при повреждении полосатого тела наблюдается атетоз - медленные червеобразные движения кистей и пальцев рук.
Дегенерация клеток этой структуры вызывает также другое заболевание - хорею, выражающуюся в судорожных подергиваниях мимических мышц и мускулатуры конечностей, которые наблюдаются в покое и при выполнении произвольных движений. Однако попытки выяснить этиологию этих явлений в экспериментах на животных не дали результатов. Разрушение хвостатого ядра у собак и кошек не приводило к возникновению гиперкинезов, характерных для описанных выше заболеваний.
Локальное электрическое раздражение некоторых участков полосатого тела вызывает у животных так называемые циркуляторные двигательные реакции, характеризующиеся поворотом головы и туловища в сторону, противоположную раздражению. Стимуляция других участков полосатого тела, напротив, приводит к торможению двигательных реакций, вызванных различными сенсорными раздражениями.
Наличие определенных расхождений между данными эксперимента и клиники, по-видимому, свидетельствует о возникновении системных нарушений механизмов регуляции движений при патологических процессах в базальных ядрах. Очевидно, эти нарушения связаны с изменениями функции не только полосатого тела, но и других структур.
В качестве примера можно рассмотреть возможный патофизиологический механизм возникновения паркинсонизма. Этот синдром связан с повреждением базальных ядер и характеризуется комплексом таких симптомов, как гипокинезия - малая подвижность и затруднения при переходе от покоя к движению; восковидная ригидность, или гипертонус, независящий от положения суставов и фазы движения; статический тремор (дрожание), наиболее выраженный в дистальных отделах конечностей.
Все эти симптомы обусловлены гиперактивностью базальных ядер, которая возникает при повреждении дофаминергического (по всей вероятности, тормозного) пути, который идет от черного вещества к полосатому телу. Таким образом, этиология паркинсонизма обусловлена дисфункцией полосатого тела и структур среднего мозга, которые функционально объединены в стриопаллидарную систему.
Для выяснения роли базальных ядер в осуществлении движений успешно используют данные микроэлектродных исследований. Эксперименты на обезьянах показали наличие корреляции между разрядами нейронов полосатого тела и медленными, направленными из стороны в сторону червеобразными движениями лапы. Как правило, разряд нейрона предшествует началу медленного движения, а при быстрых «баллистических» движениях он отсутствует. Эти факты позволяют заключить, что нейроны полосатого тела участвуют в генерации медленных движений, подвергающихся коррекции со стороны сенсорной обратной связи. Базальные ядра представляют собой один из уровней построенной по иерархическому принципу системы регуляции движений.
Получая информацию от ассоциативных зон коры, базальные ядра участвуют в создании программы целенаправленных движений с учетом доминирующей мотивации. Далее соответствующая информация от базальных ядер поступает в передний таламус, где она интегрируется с информацией, приходящей от мозжечка. Из таламических ядер импульсация достигает двигательной коры, которая отвечает за реализацию программы целенаправленного движения через посредство нижележащих стволовых и спинальных двигательных центров. Так в общих чертах можно представить место базальных ядер в целостной системе двигательных центров мозга.
Дата публикования: 2014-12-30; Прочитано: 124 | Нарушение авторского права страницы
studopedia.org — Студопедия.Орг — 2014-2018 год.(0.001 с)…
Чечевицеобразное ядро (nucl.
Базальные ядра и их функции
lentiformis) находится латерально и кпереди от таламуса. Оно имеет клиновидную форму с вершиной, обращенной к средней линии. Между задней гранью чечевицеобразного ядра и таламусом располагается задняя ножка внутренней капсулы (crus posterius capsulae internae). Передняя грань чечевицеобразного ядра внизу и спереди сращена с головкой хвостатого ядра.
Две полоски белого вещества разделяют чечевицеобразное ядро на три членика: латеральный членик – скорлупа (putamen), имеющая темную окраску, располагается с наружной стороны, а две древние части бледного шара (globus pallidus) конической формы обращены к середине.
Хвостатое ядро
Хвостатое ядро (nucl. caudatus)имеет булавовидную форму и изогнуто назад.
Передняя его часть расширена, называется головкой (caput) и располагается выше чечевицеобразного ядра, а его задняя часть — хвост (cauda) проходит сверху и латеральнее таламуса, отделяясь от него мозговыми полосками (stria medullaris). Головка хвостатого ядра участвует в образовании латеральной стенки переднего рога бокового желудочка (cornu anterius ventriculi lateralis). Хвостатое ядро состоит из малых и больших пирамидных клеток. Между чечевицеобразным и хвостатыми ядрами располагается внутренняя капсула (capsula interna).
Внутренняя капсула (capsula interna) располагается между таламусом, чечевицеобразным и хвостатым ядрами и является прослойкой белого вещества, образованной проекционными волокнами на пути к коре и от коры к нижележащим отделам ЦНС.
На горизонтальном разрезе полушария головного мозга на уровне середины таламуса внутренняя капсула имеет белую окраску и напоминает форму угла, открытого кнаружи. Внутренняя капсула разделяется на три отдела: переднюю ножку (crus anterius capsulae internae), колено (genu capsulae internae) и заднюю ножку (crus posterius capsulae internae).
Выше внутренней капсулы волокна образуют лучистый венец (corona radiata). Короткая передняя ножка капсулы образована аксонами, которые исходят из клеток коры лобной доли, и направляются в таламус (tr.
frontothalamicus), в красное ядро (tr. frontorubralis), к клеткам ядер моста (tr. frontopontinus). В колене внутренней капсулы располагается корково-ядерный путь (tr. corticonuclearis), соединяющий клетки двигательной коры с ядрами двигательных черепных нервов (III, IV, V, VII, IX, X, XI, XII). Задняя ножка внутренней капсулы несколько длиннее, чем передняя, граничит с таламусом и чечевицеобразным ядром. В передней ее части располагаются волокна, исходящие от клеток задних отделов лобной (двигательной) коры и направляющиеся к ядрам передних столбов спинного мозга.
Несколько кзади от кортико-спинального пути располагаются волокна, направляющиеся от латеральных ядер таламуса к задней центральной извилине, а также от клеток коры к ядрам таламуса. В задней ножке располагаются волокна, проходящие от коры затылочной и височной долей к ядрам моста. В заднем отделе проходят слуховые и зрительные волокна, начинающиеся от внутреннего и наружного коленчатого тел и оканчивающиеся в височной и затылочной долях.
На всем протяжении внутренней капсулы проходят поперечные волокна, которые соединяют чечевицеобразное тело с хвостатым ядром и таламусом. Веерообразно расходящиеся волокна всех проводящих путей, образующих внутреннюю капсулу, в пространстве между ней и корой полушария мозга формируют лучистый венец. Незначительные повреждения небольших участков внутренней капсулы вследствие компактности расположения волокон обусловливают тяжелые расстройства двигательных функций и потерю общей чувствительности, слуха и зрения на стороне противоположной травме.
Полосатое тело
Полосатое тело получает афферентные импульсы главным образом от таламуса, отчасти от коры; посылает эфферентные импульсы к бледному шару.
Полосатое тело рассматривают как эффекторное ядро, не имеющее самостоятельных двигательных функций, но контролирующее функции филогенетически более старого двигательного центра — паллидум а (бледного шара).
Полосатое тело регулирует и частично затормаживает безусловнорефлекторную деятельность бледного шара, т.
е. действует на него так же, как бледный шар действует на красное ядро. Полосатое тело считают высшим подкорковым регуляторно-координационным центром двигательного аппарата.
В полосатом теле, согласно экспериментальным данным, находятся также высшие вегетативные координационные центры, регулирующие обмен веществ, теплообразование и тепловыведение, сосудистые реакции.
По-видимому, в полосатом теле находятся центры, которые интегрируют, объединяют безусловнорефлекторные двигательные и вегетативные реакции в единый целостный акт поведения.
Полосатое тело оказывает влияние на органы, иннервируемые вегетативной нервной системой, через посредство своих связей с гипоталамусом. При поражениях полосатого тела у человека наблюдается атетоз — стереотипические движения конечностей, а также хорея — сильные неправильные движения, совершающиеся без всякого порядка и последовательности и захватывающие почти всю мускулатуру (“пляска святого Витта”).
И атетоз, и хорея рассматриваются как результат выпадения тормозящего влияния, которое оказывает полосатое тело на бледное ядро.
Бледный шар
Бледный шар (globus pallidus), бледное ядро, — парное образование, входящее в состав чечевицеобразного ядра, которое находится в больших полушариях и отделяется внутренней капсулой. Паллидум является двигательным ядром. При его раздражении можно получить сокращение шейных мышц, конечностей и всего туловища, преимущественно на противоположной стороне.
Бледное ядро получает импульсы по афферентным волокнам, идущим от таламуса и замыкающим таламо-паллидарную рефлекторную дугу. Бледное ядро, будучи связано эффекторно с центрами среднего и заднего мозга, регулирует и координирует их работу.
Одной из функций бледного ядра считают торможение ниже лежащих ядер, главным образом красного ядра среднего мозга, в связи с чем при повреждении бледного шара наблюдается сильное увеличение тонуса скелетной мускулатуры — гипертонус, т.
к. красное ядро освобождается от тормозящего влияния бледного шара. Таламо-гипоталамо-паллидарная система принимает участие у высших животных и человека в осуществлении сложных безусловных рефлексов — оборонительных, ориентировочных, пищевых, половых.
У человека при стимуляции бледного шара получен феномен увеличения объема кратковременной памяти почти в два раза.
Исследуя пространственно-временные соотношения между элементами речи (гласные фонемы) и регистрируемой импульсной активностью выявлена корреляция, свидетельствующая о вовлечении той или иной структуры в процесс слуховой памяти. Такие соотношения в ряде случаев удалось получить при исследовании бледного шара, дорсомедиального таламического ядра.
Миндалевидное ядро
Миндалевидное ядро (corpus amygdaloideum), или амигдалоидный комплекс представляет группу ядер и локализуется внутри переднего полюса височной доли, латеральнее перегородки продырявленного вещества.
Амигдалоидный комплекс представляет собой структуру, входящую в лимбическую систему мозга, которая характеризуется очень низким порогом возбуждения, что может способствовать развитию эпилептиформной активности.
В комплексе имеются как более крупные (пирамидные, грушевидные) и средние по размерам (мультиполярные, биполярные, канделяброобразные), так и мелкие клетки.
В амигдалоидном комплексе выделяют филогенетически более старую — кортикомедиальную — и более новую базальнолатеральную части. Группа кортикомедиальных ядер отличается низкой активностью ацетилхолинэстеразы (АХЭ) и в большей мере связана с обонятельной функцией, образуя проекции в палеокортекс. Связь с половой функцией подтверждается тем, что стимуляция этих ядер облегчает секрецию люлиберина и фоллиберина.
Нейроны базальнолатеральных ядер отличаются более высокой активностью АХЭ, дают проекцию в новую кору и полосатое тело, а также облегчают секрецию АКТГ и гормона роста. При стимуляции амигдалоидного комплекса возникают судороги, эмоционально окрашенные реакции, страх, агрессия и т. д.
Ограда
Ограда (claustrum) – тонкая прослойка серого вещества, отделенная наружной капсулой белого вещества от чечевицеобразного ядра. Ограда внизу соприкасается с ядрами переднего продырявленного вещества (substantia perforata anterior).
Предполагают участие в осуществлении глазодвигательных реакций слежения за объектом.
Предыдущая11121314151617181920212223242526Следующая
ПОСМОТРЕТЬ ЕЩЕ:
Функции базальных ядер
рис. 66). nucleus caudatus ), скорлупа (putamen ) и бледный шар (globulus pallidus claustrum ). Все эти четыре ядра называют полосатым телом (corpus striatum ).
Выделяют также стриатум (striatum nukleus lentioris

66. А — Расположение базальных ганглиев в объеме головного мозга. Базальные ганглии закрашены в красный цвет, таламус – серый цвет, а остальная часть мозга не закрашена. 1 – Бледный шар, 2 – Таламус, 3 – Скорлупа, 4 – Хвостатое ядро, 5 – Миндалевидное тело (Астапова, 2004).
У базальных ядер .
.
Возбуждающие пути

Тормозящие пути от стриатума идут к черному веществу и после переключения - к ядрам таламуса (рис.

Рис. 68. Нервные пути, секретирующие различные типы нейромедиаторов в базальных ганглиях. Ах – ацетилхолин; ГАМК – гамма-аминомасляная кислота (Гайтон, 2008)
В целом базальные ядра, имея двусторонние связи с корой большого мозга, таламусом, ядрами ствола мозга, участвуют в создании программ целенаправленных движений с учетом доминирующей мотивации. При этом нейроны стриатума оказывают тормозное влияние (медиатор - ГАМК) на нейроны черного вещества. В свою очередь, нейроны черного вещества (медиатор - дофамин) оказывают модулирующее влияние (тормозное и возбуждающее) на фоновую активность нейронов стриатума.
Функции стриатума .
Поражение
Функции бледного шара .
Ядра головного мозга и их функции
Разрушение бледного шара адинамия затрудняет осуществление имеющихся условных рефлексов и ухудшает выработку новых
Предыдущая19202122232425262728293031323334Следующая
ПОСМОТРЕТЬ ЕЩЕ:
Функции базальных ядер
Базальными ядрами называются массивные подкорковые ядра конечного мозга. Они располагаются в глубине белого вещества полушарий. К ним относятся
- хвостатое ядро (состоит из головки, тела и хвоста),
· чечевицеобразное ядро (состоит из скорлупы и бледного шара – globus pallidus – парное образование),
· ограда,
· миндалевидное тело.
Эти ядра отделены друг от друга прослойками белого вещества, образующими внутреннюю, наружную и крайнюю капсулы.
Хвостатое и чечевицеобразное ядра вместе составляют анатомическое образование — полосатое тело (corpus striatum).
Хвостатое ядро и скорлупа
Хвостатое ядро и скорлупа имеют сходное гистологическое строение.
Их нейроны относятся ко II типу клеток Гольджи, т. е. имеют короткие дендриты, тонкий аксон; их размер до 20 мк. Этих нейронов в 20 раз больше, чем нейронов Гольджи I типа, имеющих разветвленную сеть дендритов и размер около 50 мк.
Функции любых образований головного мозга определяются прежде всего их связями, которых у базальных ядер достаточно много.
Базальные ядра
Эти связи имеют четкую направленность и функциональную очерченность.
Хвостатое ядро и скорлупа получают нисходящие связи преимущественно от экстрапирамидной коры через подмозолистый пучок. Другие поля коры большого мозга также посылают большое количество аксонов к хвостатому ядру и скорлупе.
Основная часть аксонов хвостатого ядра и скорлупы идет к бледному шару, отсюда - к таламусу и только от него - к сенсорным полям.
Следовательно, между этими образованиями имеется замкнутый круг связей. Хвостатое ядро и скорлупа имеют также функциональные связи со структурами, лежащими вне этого круга: с черной субстанцией, красным ядром, люисовым телом, ядрами преддверия, мозжечком, γ-клетками спинного мозга.
Обилие и характер связей хвостатого ядра и скорлупы свидетельствуют об их участии в интегративных процессах, организации и регуляции движений, регуляции работы вегетативных органов.
Раздражение поля 8 коры большого мозга вызывает возбуждение нейронов хвостатого ядра, а поля 6 - возбуждение нейронов хвостатого ядра и скорлупы.
Одиночное раздражение сенсомоторной области коры большого мозга может вызывать возбуждение или торможение активности нейронов хвостатого ядра. Эти реакции возникают через 10-20 мс, что свидетельствует о прямых и опосредованных связях коры большого мозга с хвостатым ядром.
Медиальные ядра таламуса имеют прямые связи с хвостатым ядром, свидетельством чего служит реакция его нейронов, наступающая через 2-4 мс после раздражения таламуса.
Реакцию нейронов хвостатого ядра вызывают раздражения кожи, световые, звуковые стимулы.
Во взаимодействиях хвостатого ядра и бледного шара превалируют тормозные влияния.
Если раздражать хвостатое ядро, то большая часть нейронов бледного шара тормозится, а меньшая возбуждается. В случае повреждения хвостатого ядра у животного появляется двигательная гиперактивность.
Взаимодействие черного вещества и хвостатого ядра основано на прямых и обратных связях между ними. Установлено, что стимуляция хвостатого ядра усиливает активность нейронов черного вещества. Стимуляция черного вещества приводит к увеличению, а разрушение - к уменьшению количества дофамина в хвостатом ядре.
Установлено, что дофамин синтезируется в клетках черного вещества, а затем со скоростью 0,8 мм/ч транспортируется к синапсам нейронов хвостатого ядра. В хвостатом ядре в 1 г нервной ткани накапливается до 10 мкг дофамина, что в 6 раз больше, чем в других отделах переднего мозга, бледном шаре, в 19 раз больше, чем в мозжечке. Благодаря дофамину проявляется растормаживающий механизм взаимодействия хвостатого ядра и бледного шара.
Хвостатое ядро и бледный шар принимают участие в таких интегративных процессах, как условнорефлекторная деятельность, двигательная активность.
Это выявляется при стимуляции хвостатого ядра, скорлупы и бледного шара, деструкции и при регистрации электрической активности.
Раздражение хвостатого ядра может полностью предотвратить восприятие болевых, зрительных, слуховых и других видов стимуляции. Раздражение вентральной области хвостатого ядра снижает, а дорсальной - повышает слюноотделение.
При стимуляции хвостатого ядра удлиняются латентные периоды рефлексов, нарушается переделка условных рефлексов.
Выработка условных рефлексов на фоне стимуляции хвостатого ядра становится невозможной. Видимо, это объясняется тем, что стимуляция хвостатого ядра вызывает торможение активности коры большого мозга.
В то же время при раздражении хвостатого ядра могут появляться некоторые виды изолированных движений.
Видимо, хвостатое ядро имеет наряду с тормозящими и возбуждающие структуры.
С позиции функциональной анатомии хвостатое и чечевицеобразное ядра объединяют понятием стриопаллидарная система . Стриарная система включает в себя хвостатое ядро и скорлупу, а паллидарная – бледный шар.
Стриатум рассматривают как основное рецептивное поле стриопаллидарной системы. Здесь заканчиваются волокна из 4-х основных источников
· коры полушарий,
· зрительного бугра,
· черной субстанции,
Корковые нейроны оказывают на нейроны стриатума возбуждающее действие.
Нейроны черной субстанции оказывают на них тормозящее действие.
Аксоны нейронов стриарной системы заканчиваются на нейронах паллидум, и оказывают на них тормозящее действие.
Паллидум является выходной структурой стриопаллидарной системы.
К нему сходится основная масса эфферентных волокон.
Нейроны бледного шара оказывают на двигательные нейроны спинного мозга возбуждающее действие.
Стриопаллидарная система является центром экстрапирамидной системы. Ее основная функция – регуляция произвольных двигательных реакций. При ее участии создаются:
· оптимальная для намеченного действия поза;
· оптимальное соотношение тонуса между мышцами антагонистами и синергистами;
· плавность и соразмерность движений во времени и пространстве.
При поражении стриопаллидарной системы развивается дискинезия – нарушение двигательных актов.
Гипокинезия – бледность невыразительность движений. Усиление тормозного влияния стриарной системы на паллидарную.
Гиперкинезия (хорея) – сильные неправильные движения, совершающиеся без всякого порядка и последовательности, которые захватывают всю мускулатуру — «пляска святого Витта». Причина: выпадение тормозного влияния стриарной системы на паллидарную систему.
Ограда и миндалевидное тело входит в состав лимбической системы.
Базальные ядра обеспечивают регуляцию двигательных и вегетативных функций, участвуют в осуществлении интегративных процессов высшей нервной деятельности.
Нарушения в базальных ядрах приводит к моторным дисфункциям, таким как, замедленность движения, изменения мышечного тонуса, непроизвольные движения, тремор.
Эти нарушения фиксируются при болезни Паркинсона и болезни Хантингтона.
Бледный шар
Бледный шар (globus pallidus s. pallidum) имеет преимущественно крупные нейроны Гольджи I типа. Связи бледного шара с таламусом, скорлупой, хвостатым ядром, средним мозгом, гипоталамусом, соматосенсорной системой и др. свидетельствуют об его участии в организации простых и сложных форм поведения.
Раздражение бледного шара с помощью вживленных электродов вызывает сокращение мышц конечностей, активацию или торможение γ-мотонейронов спинного мозга.
У больных с гиперкинезами раздражение разных отделов бледного шара (в зависимости от места и частоты раздражения) увеличивало или снижало гиперкинез.
Стимуляция бледного шара в отличие от стимуляции хвостатого ядра не вызывает торможения, а провоцирует ориентировочную реакцию, движения конечностей, пищевое поведение (обнюхивание, жевание, глотание и т.д.).
Повреждение бледного шара вызывает у людей гипомимию, маскообразность лица, тремор головы, конечностей (причем этот тремор исчезает в покое, во сне и усиливается при движениях), монотонность речи.
При повреждении бледного шара наблюдается миоклония - быстрые подергивания мышц отдельных групп или отдельных мышц рук, спины, лица.
В первые часы после повреждения бледного шара в остром опыте на животных резко снижалась двигательная активность, движения характеризовались дискоординацией, отмечалось наличие незавершенных движений, при сидении - поникшая поза.
Начав движение, животное долго не могло остановиться. У человека с дисфункцией бледного шара затруднено начало движений, исчезают вспомогательные и реактивные движения при вставании, нарушаются содружественные движения рук при ходьбе, появляется симптом пропульсии: длительная подготовка к движению, затем быстрое движение и остановка. Такие циклы у больных повторяются многократно.
Ограда
Ограда (claustrum) содержит полиморфные нейроны разных типов.
Она образует связи преимущественно с корой большого мозга.
Глубокая локализация и малые размеры ограды представляют определенные трудности для ее физиологического исследования. Это ядро имеет форму узкой полоски серого вещества, расположенного под корой большого мозга в глубине белого вещества.
Стимуляция ограды вызывает ориентировочную реакцию, поворот головы в сторону раздражения, жевательные, глотательные, иногда рвотные движения.
Раздражение ограды тормозит условный рефлекс на свет, мало сказывается на условном рефлексе на звук. Стимуляция ограды во время еды тормозит процесс поедания пищи.
Известно, что толщина ограды левого полушария у человека несколько больше, чем правого; при повреждении ограды правого полушария наблюдаются расстройства речи.
Таким образом, базальные ядра головного мозга являются интегративными центрами организации моторики, эмоций, высшей нервной деятельности, причем каждая из этих функций может быть усилена или заторможена активацией отдельных образований базальных ядер.
Функции базальных ядер
Основные структуры базальных ядер (рис. 66). Базальные ядра - это хвостатое ядро (nucleus caudatus ), скорлупа (putamen ) и бледный шар (globulus pallidus ); некоторые авторы относят к базальным ядрам ограду (claustrum ).
Все эти четыре ядра называют полосатым телом (corpus striatum ). Выделяют также стриатум (striatum ) - это хвостатое ядро и скорлупа. Бледный шар и скорлупа образуют чечевицеобразное ядро (nukleus lentioris ). Стриатум и бледный шар образуют стриопаллидарную систему.
66. А — Расположение базальных ганглиев в объеме головного мозга. Базальные ганглии закрашены в красный цвет, таламус – серый цвет, а остальная часть мозга не закрашена.
1 – Бледный шар, 2 – Таламус, 3 – Скорлупа, 4 – Хвостатое ядро, 5 – Миндалевидное тело (Астапова, 2004).
Хвостатое ядро Чечевицеобразное ядро
Б – Трехмерное изображение расположения базальных ганглиев в объеме головного мозга (Гайтон, 2008)
Функциональные связи базальных ядер. У базальных ядер нет входа от спинного мозга, но есть прямой вход от коры больших полушарий .
Базальные ядра участвуют в выполнении двигательных функций, эмоциональных и познавательных (когнитивных) функций .
Возбуждающие пути идут, в основном, к стриатуму: от всех областей коры большого мозга (прямо и через таламус), от неспецифических ядер таламуса, от черного вещества (средний мозг)) (рис.
Рис. 67. Связь контура базальных ганглиев с кортикоспиномозжечковой системой для регуляции двигательной активности (Гайтон, 2008)
Сам стриатум оказывает в основном тормозное и, частично, возбуждающее влияние на бледный шар.
От бледного шара идет самый важный путь в двигательные вентральные ядра таламуса, от них возбуждающий путь идет в двигательную кору большого мозга. Часть волокон от стриатума идет в мозжечок и к центрам ствола мозга (РФ, красное ядро и далее - в спинной мозг.
Тормозящие пути от стриатума идут к черному веществу и после переключения - к ядрам таламуса (рис. 68).
68. Нервные пути, секретирующие различные типы нейромедиаторов в базальных ганглиях. Ах – ацетилхолин; ГАМК – гамма-аминомасляная кислота (Гайтон, 2008)
Двигательные функции базальных ядер. В целом базальные ядра, имея двусторонние связи с корой большого мозга, таламусом, ядрами ствола мозга, участвуют в создании программ целенаправленных движений с учетом доминирующей мотивации.
При этом нейроны стриатума оказывают тормозное влияние (медиатор - ГАМК) на нейроны черного вещества. В свою очередь, нейроны черного вещества (медиатор - дофамин) оказывают модулирующее влияние (тормозное и возбуждающее) на фоновую активность нейронов стриатума.
При нарушении дофаминергических влияний на базальные ядра наблюдаются двигательные расстройства типа паркинсонизма, при которых резко падает концентрация дофамина в обоих ядрах стриатума. Наиболее важные функции базальных ядер выполняют стриатум и бледный шар.
Функции стриатума .
Участвует в осуществлении поворота головы и туловища и ходьбы по кругу , которые входят в структуру ориентировочного поведения. Поражение хвостатого ядра при заболеваниях и при разрушении в эксперименте ведет к насильственным, избыточным движениям (гиперкинезы: хорея и атетоз).
Функции бледного шара .
Оказывает модулирующее влияние на двигательную кору, мозжечок, РФ, красное ядро. При стимуляции бледного шара у животных преобладают элементарные двигательные реакции в виде сокращения мышц конечностей, шеи и лица, активация пищевого поведения.
Разрушение бледного шара сопровождается снижением двигательной активности - возникает адинамия (бледность двигательных реакций), а также ему (разрушению) сопутствует развитие сонливости, «эмоциональной тупости», что затрудняет осуществление имеющихся условных рефлексов и ухудшает выработку новых (ухудшает кратковременную память).
Движение и мышление – это те качества, которые позволяют человеку полноценно жить и развиваться.
Даже незначительные нарушения в структурах мозга могут привести к существенным изменениям или полной утрате этих способностей.
Ответственными за эти важнейшие жизненные процессы являются группы нервных клеток головного мозга, называемые базальными ядрами.
Что нужно знать о базальных ядрах
Большие полушария мозга человека снаружи представляют собой кору, образованную серым веществом, а внутри – подкорку из белого вещества. Базальные ядра (ганглии, узлы), которые также называют центральными, или подкорковыми – это сосредоточения серого вещества в белом веществе подкорки.
Базальные ганглии расположены в основании головного мозга, что и объясняет их название, снаружи от таламуса (зрительного бугра). Это парные образования, которые симметрично представлены в обоих полушариях мозга. При помощи нервных отростков они в двустороннем порядке взаимодействуют с различными областями центральной нервной системы.
Основная роль подкорковых узлов заключается в организации двигательной функции и различных аспектов высшей нервной деятельности. Патологии, которые возникают в их строении, влияют на работу других частей центральной нервной системы, вызывая проблемы с речью, координацией движений, памятью, рефлексами.
Особенности строения базальных узлов
Базальные ганглии находятся в лобных и частично височных долях конечного мозга. Это скопления тел нейронов, которые образуют группы серого вещества. Окружающее их белое вещество представлено отростками нервных клеток и формирует прослойки, разделяющие отдельные базальные ядра и прочие мозговые структурно-функциональные элементы.
К базальным узлам причисляются:
- полосатое тело;
- ограда;
- миндалевидное тело.
На анатомических срезах полосатое тело выглядит как перемежающиеся слои серого и белого вещества. В его составе выделяют хвостатое и чечевицеобразное ядра. Первое расположено кпереди от зрительного бугра. Истончаясь, хвостатое ядро переходит в миндалевидное тело. Чечевицеобразное ядро находится латеральнее зрительного бугра и хвостатого ядра. Оно соединяется с ними тонкими перемычками нейронов.
Ограда – это узкая полоска нейронов. Она расположена между чечевицеобразным ядром и островковой корой мозга. От этих структур ее отделяют тонкие слои белого вещества. Миндалевидное тело по форме напоминает миндалину и находится в височных долях конечного мозга. В его составе различают несколько самостоятельных элементов.
Данная классификация построена на особенностях строения и расположения ганглиев на анатомическом срезе мозга. Существует также функциональная классификация, согласно которой ученые причисляют к базальным узлам только полосатое тело и некоторые ганглии промежуточного и среднего мозга. Эти структуры в комплексе обеспечивают двигательные функции человека и отдельные аспекты поведения, отвечающие за мотивацию.
Анатомия и физиология базальных ядер
Хотя все базальные ганглии являются скоплением серого вещества, они имеют свои сложные структурные особенности. Чтобы понять, какую роль играет тот или иной базальный центр в работе организма, необходимо подробнее рассмотреть его строение и расположение.
Хвостатое ядро
Этот подкорковый узел расположен в лобных долях полушарий мозга. Его подразделяют на несколько отделов: утолщенную крупную головку, сужающееся тело и тонкий длинный хвост. Хвостатое ядро сильно вытянуто и изогнуто. Ганглий состоит большей частью из микронейронов (до 20 мк) с короткими тонкими отростками. Около 5% от общей клеточной массы подкоркового узла составляют более крупные нервные клетки (до 50 мк) с сильноветвящимися дендритами.
Данный ганглий взаимодействует с участками коры, таламусом и узлами промежуточного и среднего мозга. Он исполняет роль связующего звена между этими мозговыми структурами, постоянно передавая нейронные импульсы от коры мозга к другим его отделам и обратно. Он многофункционален, но особенно значительна его роль в поддержании активности нервной системы, регулирующей деятельность внутренних органов.
Чечевицеобразное ядро
Этот базальный узел своей формой имеет схожесть с семенем чечевицы. Он также располагается в лобных отделах больших полушарий. При разрезе головного мозга во фронтальной плоскости данная структура представляет собой треугольник, вершина которого направлена внутрь. Белым веществом этот ганглий подразделяется на скорлупу и два слоя бледного шара. Скорлупа темная и расположена наружно по отношению к светлым прослойкам бледного шара. Нейронный состав скорлупы аналогичен хвостатому ядру, а вот бледный шар представлен в основном крупными клетками с небольшими вкраплениями микронейронов.
Эволюционно бледный шар признан самым древним образованием среди прочих базальных узлов. Скорлупа, бледный шар и хвостатое ядро составляют стриопаллидарную систему, являющуюся частью экстрапирамидной. Основная функция этой системы – регуляция произвольных движений. Анатомически она связана с множеством корковых полей больших полушарий.
Ограда
Слегка изогнутая истонченная пластина серого вещества, которая разделает скорлупу и островковую долю конечного мозга, получила название ограды. Белое вещество вокруг нее образует две капсулы: наружную и «самую наружную». Эти капсулы отделяют ограду от соседних структур из серого вещества. Ограда прилегает к внутреннему слою новой коры головного мозга.
Толщина ограды варьирует от долей миллиметра до нескольких миллиметров. На всем протяжении она состоит из нейронов различной формы. Нервными путями ограда связана с центрами коры больших полушарий, гиппокампом, миндалевидным и частично полосатым телами. Отдельные ученые считают ограду продолжением коры мозга или же вносят ее в состав лимбической системы.
Миндалевидное тело
Данный ганглий представляет группу клеток серого вещества, сосредоточенных под скорлупой. Миндалевидное тело состоит из нескольких образований: ядер коры, срединного и центрального ядра, базолатерального комплекса, интерстициальных клеток. Оно связано нервной передачей с гипоталамусом, таламусом, органами чувств, ядрами черепно-мозговых нервов, центром обоняния и многими другими образованиями. Иногда миндалевидное тело причисляют к лимбической системе, которая отвечает за деятельность внутренних органов, эмоции, обоняние, сон и бодрствование, обучение и т.д.
Важность подкорковых узлов для организма
Функции базальных узлов определяются их взаимодействием с другими областями центральной нервной системы. Они формируют нейронные петли, соединяющие таламус и важнейшие зоны коры полушарий мозга: моторную, соматосенсорную и лобную. Кроме того, подкорковые узлы связаны между собой и с некоторыми областями ствола мозга.
Хвостатое ядро и скорлупа выполняют следующие функции:
- контроль направления, силы и амплитуды движений;
- аналитическая деятельность, обучение, мышление, память, коммуникация;
- управление движением глаз, рта, лица;
- поддержание работы внутренних органов;
- условнорефлекторная деятельность;
- восприятие сигналов органов чувств;
- контроль мышечного тонуса.
К специфичным функциям скорлупы относят дыхательные движения, выработку слюны и другие аспекты пищевого поведения, обеспечение трофики кожи и внутренних органов.
Функции бледного шара:
- развитие ориентировочной реакции;
- контроль движения рук и ног;
- пищевое поведение;
- мимика;
- проявление эмоций;
- обеспечение вспомогательных движений, координационных способностей.
К функциям ограды и миндалевидного тела относятся:
- речь;
- пищевое поведение;
- эмоциональная и долгосрочная память;
- развитие поведенческих реакций (страх, агрессия, тревожность и др.);
- обеспечение социальной интеграции.
Таким образом, размер и состояние отдельных базальных узлов влияет на эмоциональное поведение, произвольные и непроизвольные движения человека, а также высшую нервную деятельность.
Заболевания базальных узлов и их симптомы
Нарушение нормального функционирования базальных ядер может быть вызвано инфекцией, травмой, генетической предрасположенностью, врожденными аномалиями, сбоем в обмене веществ.
Симптомы патологии иногда проявляются постепенно, незаметно для пациента.
Следует обратить внимание на такие признаки:
- общее ухудшение самочувствия, слабость;
- нарушение тонуса мускулатуры, ограниченность движений;
- возникновение произвольных движений;
- тремор;
- нарушение координации движений;
- возникновение непривычных для пациента поз;
- обеднение мимики;
- нарушение памяти, помутнение сознания.
Патологии базальных ганглиев могут проявляться рядом заболеваний:
- Функциональная дефицитарность. Преимущественно наследственное заболевание, проявляющееся в детском возрасте. Основные симптомы: неуправляемость, невнимательность, энурез до 10–12 лет, неадекватное поведение, нечеткость движений, странные позы.
- Киста. Злокачественные образования без своевременного врачебного вмешательства приводят к инвалидности и смерти.
- Корковый паралич. Основные симптомы: непроизвольные гримасы, нарушение мимики, судороги, хаотичные медленные движения.
- Болезнь Паркинсона. Основные симптомы: тремор конечностей и тела, обеднение двигательной активности.
- Болезнь Хантингтона. Генетическая патология, прогрессирующая постепенно. Основные симптомы: спонтанные неконтролируемые движения, нарушение координации, снижение умственных способностей, депрессия.
- . Основные симптомы: замедление и обеднение речи, апатия, неадекватное поведение, ухудшение памяти, внимания, мышления.
Некоторые функции базальных ганглиев и особенности их взаимодействия с другими структурами мозга до сих пор не установлены. Неврологи продолжают изучать эти подкорковые центры, ведь их роль в поддержании нормальной жизнедеятельности организма человека бесспорна.
БАЗАЛЬНЫЕ ЯДРА [позднелатинский basalis относящийся к основанию; синоним: центральные узлы, подкорковые ядра (nuclei subcorticales )] - скопления серого вещества в толще больших полушарий головного мозга, участвующие в коррекции программы сложного двигательного акта и формировании эмоционально-аффективных реакций.
Первые сведения по морфологии базальные ядра встречаются в работах Бурдаха (К. F. Burdach), 1819; И. П. Лебедева, 1873; Антона (Anton), 1895; Капперса (С. A. Kappers), 1908, и др. Большой вклад в изучение базальных ядер внесли анатомические и клинико-морфологические исследования С. Фогт и О. Фогта (С. Vogt, О. Vogt), 1920; М. О. Гуревич, 1930; Фуа и Николеско (Foix, Nicolesco), 1925; E. Κ. Сеппа, 1949; T. А. Леонтовича, 1952, 1954; Η. П. Бехтеревой, 1963; Э. И. Канделя, 1961; Л. А. Кукуева, 1968, и др.
Базальные ядра наряду с расположенной по поверхности полушарий корой мозга (cortex cerebri) составляют клеточное вещество конечного мозга (telencephalon). В отличие от коры, имеющей строение экранных центров (характеризующееся определенными цитоархитектоническими признаками: четким выделением слоев, вертикальной ориентацией большинства нейронов, их дифференцировкой по форме и величине в зависимости от их положения в разных слоях), базальные ядра имеют строение ядерных центров, где подобная структурная организация отсутствует. Нередко эти ядра называют подкоркой. К ним относятся: хвостатое ядро (nucleus caudatus), чечевицеобразное ядро (nucleus lentiformis, s. nucleus lenticularis), ограда (claustrum) и миндалевидное тело (corpus amygdaloideum). К базальным ядрам относят также базальный комплекс ядер, расположенный между передним продырявленным веществом (substantia perforata anterior) и передней частью бледного шара (globus pallidus), принадлежащий септальной области (см.).
Сравнительная анатомия
Исследования развития базальных ядер в фило и онтогенезе показали, что хвостатое ядро и скорлупа чечевицеобразного ядра (putamen) развиваются из ганглиозного бугра, расположенного на нижней стенке бокового желудочка. Они представляют единую клеточную массу, которая у высших позвоночных разделяется волокнами передней ножки внутренней капсулы (crus anterior capsulae internae). Ввиду общности происхождения и сохраняющегося в течение всей жизни соединения головки хвостатого ядра и переднего отдела скорлупы полосками серого вещества, чередующимися с белыми пучками волокон внутренней капсулы, хвостатое ядро и скорлупу объединяют под названием «полосатое тело» (corpus striatum), или «стриатум» (striatum). Так как полосатое тело является филогенетически более поздним образованием, чем медиально расположенная часть чечевицеобразного ядра - бледный шар, состоящий из наружного и внутреннего члеников, его называют «неостриатум» (neostriatum), а бледный шар - «палеостриатум» (paleostriatum). Последний в наст, время выделяют в особую морфологическую единицу под названием «паллидум» (pallidum).
Исследования Л. А. Кукуева (1968) показывают, что наружный и внутренний членики бледного шара имеют различное происхождение. Наружный членик, как и скорлупа, развивается из ганглиозного бугра конечного мозга; внутренний членик - из промежуточного мозга и гомологичен энтопедункулярному ядру субприматов (расположено у них в мозге над зрительным трактом, то есть его топография сходна с топографией внутреннего членика бледного шара на ранних стадиях развития зародыша человека). В процессе как филогенетического, так и онтогенетического развития происходит перемещение внутреннего членика по направлению к наружному, в результате чего они сближаются.
Базальные ядра различно представлены в мозге разных классов позвоночных животных. Так, у рыб и амфибий базальные ядра представлены лишь бледным шаром, хвостатое ядро и скорлупа появляются впервые у рептилий, особенно хорошо они развиты у птиц. У млекопитающих (хищных и грызунов) бледный шар представлен единым образованием, у человека он состоит из двух члеников, разделенных прослойкой белого вещества. Размеры полосатого тела уменьшаются по мере развития головного мозга в филогенезе. Из млекопитающих у низших насекомоядных оно составляет 8% от величины всего конечного мозга, у тупайя и полуобезьян -7%, а у обезьян-6%.
В онтогенезе полосатое тело можно дифференцировать в начале 2-го месяца эмбрионального развития. На 3-м месяце развития головка хвостатого ядра вдается в полость бокового желудочка. Латеральнее хвостатого ядра формируется скорлупа, которая вначале нерезко отграничена от остальных частей полушария. Миндалевидное тело занимает особое положение среди базальных ядер; на ранних стадиях эмбрионального развития оно отделяется от полосатого тела, цитологическая дифференцировка происходит в нем позднее, чем в бледном шаре, однако несколько раньше, чем в полосатом теле. Исходя из онто- и филогенетического развития, нельзя его также рассматривать и как измененную, утолщенную часть коры височной доли или как результат ее погружения внутрь и отшнуровывания. При исследовании миндалевидного тела в сравнительно-анатомическом аспекте было выявлено заметное уменьшение его размеров у млекопитающих - от низших насекомоядных, где оно вместе с палеокортексом составляет 31% от всей величины конечного мозга, до человека, в головном мозге которого миндалевидное тело составляет лишь 4% от всей массы конечного мозга. Исследования развития ограды в онто- и филогенезе (И. Н. Филимонов) показали, что ее нельзя считать производным корковой пластинки или связывать по происхождению с полосатым телом. Она представляет собой промежуточное образование между этими основными клеточными массами конечного мозга.
Анатомия
Хвостатое ядро имеет грушевидную форму; его передняя часть утолщена и носит название головки хвостатого ядра (caput nuclei caudati). Она расположена в переднем отделе полушария и выступает в передний рог бокового желудочка (cornu anterius ventriculi lateralis), образуя его стенку снизу и латерально. Кзади от головки хвостатое ядро суживается и этот отдел его называется телом хвостатого ядра (corpus nuclei caudati). Тело хвостатого ядра ограничивает с латеральной стороны центральную часть бокового желудочка (pars centralis ventriculi lateralis) и описывает полукруг над зрительным бугром (thalamus) и чечевицеобразным ядром. Истонченный задний отдел хвостатого ядра, образующий часть крыши нижнего рога бокового желудочка (cornu inferius ventriculi lateralis), образует хвост хвостатого ядра (cauda nuclei caudati). Латеральная поверхность хвостатого ядра прилежит к внутренней капсуле (capsula interna), его медиальный край примыкает к конечной полоске (stria terminalis).
Чечевицеобразное ядро имеет форму клина, основание которого направлено латерально, а верхушка медиально и вниз, примыкая к подбугорной области. Оно лежит латерально и несколько ниже (вентральнее) от хвостатого ядра и зрительного бугра, от которых отделено внутренней капсулой. Спереди и вентрально чечевицеобразное ядро соединяется с головкой хвостатого ядра тонкими полосками серого вещества. Латеральная его поверхность несколько выпуклая и располагается вертикально, гранича с наружной капсулой (capsula externa), которая представляет собой тонкую белую мозговую пластинку, ограниченную латерально серым веществом - оградой (claustrum). Вентральная поверхность чечевицеобразного ядра лежит горизонтально и в средней своей части соединяется с корой в области переднего продырявленного вещества. Две тонкие мозговые пластинки, медиальная и латеральная (laminae medullares medialis et lateralis), разделяют его на три части: наружная часть, более темно окрашенная, называется скорлупой, две другие представляют собой более слабо окрашенные наружный и внутренний членики бледного шара. Ограда является узкой пластинкой серого вещества, которая расположена латеральнее чечевицеобразного ядра и отделена от него наружной капсулой. От коры островка (insula) ограда отделена слоем белого вещества, образующим внешнюю капсулу (capsula extrema).
Миндалевидное тело - это комплекс ядер, расположенных в области крючка парагиппокампальной извилины (uncus gyri parahippocampalis), хорошо дифференцированных и отличающихся друг от друга цитологически и цитоархитектонически (см. Амигдалоидная область).
Гистология
Хвостатое ядро и скорлупа сходны по гистологическому строению. Серое вещество этих ядер состоит из двух видов клеточных элементов: малых и крупных клеток. Малые клетки, величиной до 15-20 мкм, имеющие короткие дендриты и тонкие аксоны, обладают нежной зернистостью и большим ядром с ядрышком. Крупные клетки, размерами до 50 мкм, в основном треугольные и полигональные, ядро их часто расположено эксцентрично, в протоплазме имеются хроматиновые зернышки и по соседству с ядром большое количество желтого липоидного пигмента. Эти клетки в норме окружены сателлитами. Отношение крупных клеток к малым в хвостатом ядре и скорлупе в среднем составляет 1: 20. Как малые, так и крупные клетки имеют длинные аксоны, прослеживаемые к другим глубоким структурам мозга.
Рис. 1. Схема главных связей экстрапирамидальной системы (по С. и О. Фогт): 7 -cortex prefrontalis; 2 - tractus frontothalamicus; 3 - nucleus caudatus; 4 - thalamus; 5 -nucleus medialis thalami; 6 и 25 - nucleus ventralis thalami; 7 -nucleus campi Forell (BNA); 8 - nucleus subthalamicus; 9 -decussatio Foreli (BNA); 10 - nucleus ruber; 11 - substantia nigra; 12 - comissura post.; 13 - nucleus Darkschewitschi; 14 - nucleus interstitialis; 15 - pedunculi cerebelli superiores (tractus cerebellotegmentalls); 16 - cerebellum; 17 - nucleus dentatus; 18 - pedunculi cerebelli medii; 19 - nucleus vestibularis sup.; 20 - canalis semicirculatis; 21 - nucleus vestibularis lat.; 22 - fasciculus longitudinalis medius; 23 - fasciculus rubrospinalis; 24 - crus cerebri; 26 - globus pallidus; 27 - putamen; 28 - area gigantopyramidalis; 29 - capsula interna.


Определенные взаимоотношения между клеточными элементами и волокнами позволили Фогту (О. Vogt) указать на сходство строения полосатого тела с корой. В хвостатом ядре под эпендимой располагается зона, бедная волокнами; наружная часть этой зоны бедна ганглиозными клетками, внутренняя ими более богата. Глубже находится слой тангенциальных волокон, содержащий небольшое количество ганглиозных клеток. На основании этого Фогтом была разработана схема структурнофункциональной организации полосатого тела (цветной рис. 1): стриопетальные волокна оканчиваются на малых клетках, тесно связанных между собой и с большими клетками, от которых уже начинаются стриофугальные волокна. В малых клетках фибриллы не дифференцированы, в больших - распределены пучками. Миелиновых волокон в полосатом теле мало, большая часть их возникает в самом полосатом теле и служит для соединения с паллидумом; между пучками миелиновых волокон располагается густая сеть безмиелиновых. Богатая сеть нейроглии окружает нервные клетки и нервные волокна. В паллидуме имеются клетки только очень крупные, различной формы - пирамидные, веретенообразные, муль-типолярные с длинными дендритами (цветные рис. 2 и 3). В протоплазме много хроматофильных глыбок. Поверхность клеток покрыта петлеобразными концевыми тельцами - окончаниями безмиелиновых волокон, окружающих клетки и миелиновые волокна. Миелиновых волокон много больше, чем серого вещества; этим объясняется бледный цвет ядра.
Кровоснабжение базальных ядер осуществляется главным образом от средней мозговой артерии (a. cerebri media), ветвями, идущими к полосатому телу (rr. striati). Ветви передней мозговой артерии (a. cerebri anterior) также принимают участие в кровоснабжении базальных ядер. Все базальные ядра, особенно полосатое тело, очень богаты капиллярами; распределение капилляров в полосатом теле напоминает распределение в коре; при поражениях сосудов мозга в полосатом теле особенно часто появляются очаги размягчения.
Связи базальных ядер
Афферентные волокна полосатое тело получает от зрительного бугра, от ядер подбугорья, окружающих III желудочек, из покрышки среднего мозга (tegmentum mesencepnali) и от черного вещества (substantia nigra). Эти волокна заканчиваются около малых клеток полосатого тела, от которых аксоны преимущественно идут к большим клеткам, а уже от этих последних волокна идут в паллидум в составе стрио-паллидарного пучка (fasciculus striopallidalis). Волокна хвостатого ядра пересекают внутреннюю капсулу, входят в скорлупу, а затем, пронизывая мозговую пластинку, проникают в паллидум. Из скорлупы, от ее крупных клеток, волокна через мозговую пластинку также входят в паллидум. Последний является главным местом, куда направляются волокна от хвостатого ядра и скорлупы. Некоторые авторы не отрицают возможности существования длинных волокон, идущих непосредственно из скорлупы в ствол, не прерываясь в паллидуме. Афферентные волокна, направляющиеся в паллидум, состоят из волокон, идущих: 1) непосредственно от коры; 2) из коры через зрительный бугор; 3) от полосатого тела; 4) от центрального серого вещества (substantia grisea centralis) промежуточного мозга; 5) от крыши (tectum) и покрышки (tegmentum) среднего мозга; 6) от черного вещества.
Эфферентные волокна базальных ядер отходят от бледного шара. Главный пучок, выходящий из него,- лентикулярная петля (ansa lenticularis); ее волокна начинаются в хвостатом ядре, принимают участие в образовании мозговых пластинок (laminae medullares). Петля прерывается в бледном шаре. Волокна, выходящие из бледного шара, пересекают внутреннюю капсулу; на границе с ножками мозга в подбугорье они рассыпаются веерообразно и заканчиваются в переднем и латеральном ядрах зрительного бугра, в подбугорье (hypothalamus), черном веществе, подбугорном ядре (nucleus subthalamicus) и красном ядре (nucleus ruber). Часть волокон идет в составе переднего перекреста покрышки (decussatio tegmentalis anterior) на противоположную сторону, где заканчивается в одноименных образованиях. Другой пучок, выходящий из бледного шара, - лентикулярный пучок (fasciculus lenticularis). Этот пучок располагается под zona incerta, включает волокна, идущие к подбугорному ядру (вокруг которого образуют сумку), к зрительному бугру, красному ядру, ядру нижней оливы (nucleus olivaris), сетчатому веществу (formatio reticularis), четверохолмию , перивентрикулярным ядрам. Часть волокон через передний перекрест покрышки переходит на противоположную сторону и заканчивается в тех же образованиях. Описаны пути, соединяющие полосатое тело с областью воронки (infundibulum) и располагающиеся над zona incerta. От красного ядра, четверохолмия начинаются периферические Экстрапирамидные волокна (tractus rubrospinalis, tractus tectospinalis). Точных данных о связи ограды и миндалевидного тела пока нет. В литературе имеются указания на связь у животных ограды с волокнами из наружной сумки, происходящими из пириформной области, на ее связь с миндалевидным телом противоположной области и вентральной областью промежуточного мозга. Установлено также, что ограда связана с корой островка. Связи миндалевидного тела - см. Амигдалоидная область .
Физиология базальных ядер

Рис. Основные афферентные и эфферентные связи (обозначены стрелками) базальных ядер с другими системами мозга (I, II, IV - по Бьюси; III - по Глису): I - связи из моторной и премоторной зон (поля 4, 4S, 6,8, 24) коры мозга к хвостатому ядру и скорлупе; II - связи базальных ядер с ядрами зрительного бугра; III - связи между отдельными базальными ядрами и между базальными ядрами и моторной и премоторной зонами коры; IV - связи базальных ядер с черной субстанцией и красным ядром. С. N. (С - по Глису) - nuci, caudatus; V. А. (Nva - по Глису) - nuci, ventralis ant. thalami; V. L.- nuci, lateralis thalami; V. P.- nuci, ventralis post, thalami; С. М.- nuci, medialis thalami; R. N.- nuci, ruber; S. N.- substantia nigra; C. e.- corpus callosum; F - fornix; Na - nuci. ant. thalami; Tr. o.- tractus opticus; P - putamen; Pi - globus pallidus (внутренний членик); Pe - globus pallidus (наружный членик); Ca - comissura ant.; Th - thalamus; G. P.- globus pallidus; H.- hypothalamus; С. S.- sulcus centralis.
На низших ступенях эволюции (у рыб, рептилий, птиц) базальные ядра являются высшими центрами координации сложного поведения. У человека и высших животных (приматов) сложную интегративную деятельность осуществляет кора больших полушарий, однако роль базальных ядер не уменьшается, а лишь видоизменяется (Е. К. Сепп, 1959).
На ранних стадиях постнатального онтогенеза основная двигательная функция новорожденного - непроизвольные хаотические движения - осуществляется в основном за счет паллидума. С развитием стриатума в более поздние сроки постнатального онтогенеза отмечаются эмоциональные проявления (улыбка) и усложняются статокинетические и тонические функции (ребенок удерживает головку, выполняет содружественные движения). При рассмотрении физиологической роли базальных ядер необходимо исходить из особенностей связей этих ядер с другими отделами головного мозга (Е. П. Кононова, 1959; И. Н. Филимонов, 1959; О. Загер, 1962). Базальные ядра характеризуются богатством афферентных и эфферентных связей с моторными зонами коры головного мозга (рис., /), с ядрами зрительного бугра (рис., II), между базальными ядрами (рис., III), с ядрами среднего мозга (рис., IV), а также с подбугорьем, образованиями лимбической системы и мозжечком. Важное значение для понимания физиологии базальных ядер имеет учет обратных связей, идущих от них в кору головного мозга. Такой широкий спектр связей обусловливает сложность функционального значения базальных ядер (объединяемых в стрио-паллидарную систему) в различных нейрофизиологических и психофизиологических процессах (В. А. Черкес, 1963; Е. Ю. Ривина, 1968; Η. П. Бехтерева, 1971). Установлено участие базальных ядер в следующих нейрофизиологических функциях: а) сложные двигательные акты; б) вегетативные функции; в) безусловные рефлексы; г) сенсорные процессы; д) условнорефлекторные механизмы; е) психофизиологические процессы (эмоции). Роль базальных ядер в осуществлении сложных двигательных актов заключается в том, что они обусловливают миостатические реакции, оптимальное перераспределение мышечного тонуса (благодаря модулирующим влияниям на нижележащие структуры центральной нервной системы, определяющие регуляцию движений).
Так, изучение функции паллидума, проведенное в условиях хронического опыта, позволило установить его важную роль в протекании сложных безусловных рефлексов различной биологической, направленности -половых, пищевых, оборонительных и др.
Методом прямой электростимуляции паллидума показана легкость воспроизведения моторных и биоэлектрических проявлений эпилептиформных реакций тонического типа. Среди важнейших функций хвостатого ядра и скорлупы следует отметить их тормозящее влияние на паллидум [Тилни и Рили (F. Tilney, H. А. Riley), 1921; Пейпс (J. W. Papez), 1942; А. М. Гринштейн, 1946, и др.]. Эффекты выключения неостриатума (полосатое тело) отражаются на функциональной активности паллидарных и среднемозговых центров (черная субстанция, ретикулярная формация ствола). Происходит их растормаживание, что сопровождается изменением мышечного тонуса и появлением гиперкинезов (см.). Многочисленные исследования влияния хвостатого ядра на условнорефлекторную деятельность и на целенаправленные движения свидетельствуют как о тормозящем, так и об облегчающем характере этих влияний, что привело к заключению о наличии двух восходящих активирующих систем: неостриарной и ретикулярной; неостриарная осуществляет влияния на кору больших полушарий как прямо, так и опосредованно, через ядра зрительного бугра. В базальных ядрах обнаружены явления конвергенции звуковых, зрительных, проприоцептивных импульсов. По-видимому, базальные ядра являются передаточной инстанцией импульсов из ретикулярной формации в кору больших полушарий. Этим объясняются явления дезориентации, хаотической двигательной активности на фоне стимуляции хвостатого ядра и скорлупы. Важное значение полосатое тело имеет в регуляции вегетативных компонентов сложных поведенческих реакций. Раздражение неостриатума сопровождается эмоционально выразительными реакциями (мимические реакции, повышенная двигательная активность). При лечении больных в нейрохирургических клиниках, проводимом с помощью вживленных на длительный срок электродов, показано угнетающее влияние стимуляции хвостатого ядра на выполнение интеллектуальной, речевой деятельности, на состояние памяти (Η. П. Бехтерева, 1971, и др.). Большое значение придают базальные ядра в механизме развития гиперкинезов. При разрушении паллидума или его патологии наблюдается проявление гипертонии мышц, ригидность, гиперкинез. Однако установлено, что развитие гиперкинезов есть результат выпадения функции не отдельного базального ядра, а сопряжено с нарушением функций вентро-медиальных ядер зрительного бугра и центров среднего мозга, регулирующих тонус (В. А. Черкес, 1963; Η. П. Бехтерева, 1965, 1971).
Данные нейрофизиологических и клинико-неврологических исследований функций базальных ядер позволяют заключить, что их физиологическое значение необходимо рассматривать в связи с другими системами мозга. Хартманн и Монаков (Н. Hartmann, К. Monakow, 1960) показали, что во время сложного двигательного акта базальные ядра объединяются непрерывным потоком импульсов, которые распространяются по определенным нейронным кругам: а) зрительный бугор - полосатое тело - зрительный бугор; б) зрительный бугор - кора полушарий - полосатое тело - бледный шар - зрительный бугор.
Функциональные взаимоотношения между базальными ядрами еще до конца не выяснены. Электрофизиологические исследования показали, что контроль полосатого тела над бледным шаром не является только тормозным. В острых опытах на кошках было выявлено также и облегчающее влияние хвостатого ядра на нейронную активность бледного шара, о чем свидетельствует учащение потенциалов действия отдельных элементов бледного шара под влиянием раздражения головки хвостатого ядра.
Изучение вызванных потенциалов в базальных ядрах показало возможность конвергенции возбуждений от различных сенсорных каналов на одном и том же нейроне [Сегундо и Макне (I. P. Segundo, X. Machne), 1956; Альб-Фессар с соавторами (D. Albe-Fessard) с соавт., 1960], и, по их мнению, ни в одной из нейронных групп базальных ядер не представлена соматотопическая локализация.
Большой удельный вес афферентных морфо-функциональных связей позволяет считать, что физиологическая роль базальных ядер не исчерпывается двигательной сферой. Учитывая большое значение обратных связей и тесное взаимодействие базальных ядер с другими системами мозга, можно прийти к заключению, что роль базальных ядер заключается в сличении различных афферентных воздействий для выполнения конечной двигательной задачи. Исходя из концепции П. К. Анохина о функциональной системе (1968), можно считать, что базальные ядра участвуют в формировании афферентного синтеза, в коррекции программы сложного двигательного акта и в оценке результатов действия. Кроме того, функциональное состояние базальных ядер отражается и на других функциях головного мозга, особенно при формировании эмоционально-аффективных реакций.
Библиогр. Анохин П. К. Биология и нейрофизиология условного рефлекса, М., 1968, библиогр.; Беритов И. С. Нервные механизмы поведения высших позвоночных животных, М., 1961, библиогр.; Бехтерева Η. П. Нейрофизиологические аспекты психической деятельности человека, Л., 1971, библиогр.; Беляев Ф. П. Подкорковые механизмы сложных двигательных рефлексов, Д., 1965, библиогр.; Гранит Р. Электрофизио-логическое исследование рецепции, пер. с англ., М., 1957, библиогр.; К о г а н А. Б. Электрофизиологическое исследование центральных механизмов некоторых сложных рефлексов, М., 1949, библиогр.; Рожанский Н. А. Очерки по физиологии нервной системы, JI., 1957, библиогр.; Сепп Е. К. История развития нервной системы позвоночных. М., 1959, библиогр.; Суворов Η. Ф. Центральные механизмы сосудистых нарушений, JI., 1967, библиогр.; Филимонов И. Н. Филогенез и онтогенез нервной системы, Многотомн. руководство по неврол., под ред. Н. И. Гращенкова, т. 1, кн. 1, с. 9, М., 1959; Черкес В. А. Очерки по физиологии базальных ганглиев головного мозга, Киев, 1963, библиогр.; А 1 b е-Fessard D., Oswaldo-Cruz E. a. Rocha-M iranda С. Activity 6voqu6es dans le noyau caude du chat en rSponse h des types divers d’aff6rences, Electroenceph. clin. Neurophysiol., v. 12, p. 405, 1960; B u с у Р. С. The basal ganglia, the thalamus and hypothalamus, в кн.: Physiol, basis med. pract., ed. by С. H. Best, p. 144, Baltimore, 1966, bibliogr.; Clara M. Das Nervensystem des Menschen, Lpz., 1959, Bibliogr.; The diseases of the basal ganglia, ed. by T. J. Putnam a. o., Baltimore, 1942, bibliogr.
H. H. Боголепов, E. П. Кононова; Ф. П. Ведяев (физ.).
Базальные (подкорковые) ядра располагаются под белым веществом внутри переднего мозга, преимущественно в лобных долях. У млекопитающих к базальным ганглиям относятся сильно вытянутое в длину и изогнутое хвостатое ядро и заложенное в толще белого вещества чечевицеобразное ядро. Двумя белыми пластинками оно подразделяется на три части: наиболее крупную, лежащую латерально скорлупу и бледный шар, состоящий из внутреннего и внешнего отделов. Они формируют так называемую стриопаллидарную систему, которая по филогенетическим и функциональным критериям разделяется на древний палеостриатум и неостриатум. Палеостриатум представлен бледным шаром, а неостриатум, состоит из хвостатого ядра и скорлупы, которые объединяются под названием полосатого тела или стриатума. А объединяют их под общим названием «полосатое тело», в связи с тем, что скопление нервных клеток, образующих серое вещество, чередуются с прослойками белого вещества. (Ноздрачева А.Д., 1991)
Базальные ганглии головного мозга человека включает в себя также ограду. Это ядро имеет форму узкой полоски серого вещества. (Покровский, 1997) Медиально она граничит с наружной капсулой, латерально - с капсулой экстрема.
Нейронная организация
Хвостатое ядро и скорлупа имеют сходную нейронную организацию. Они содержат главным образом мелкие нейроны с короткими дендритами и тонкими аксонами, их размер до 20 мк. Кроме мелких, имеется небольшое число (5% от общего состава) относительно крупных нейронов, имеющих разветвленную сеть дендритов и размер около 50 мк.
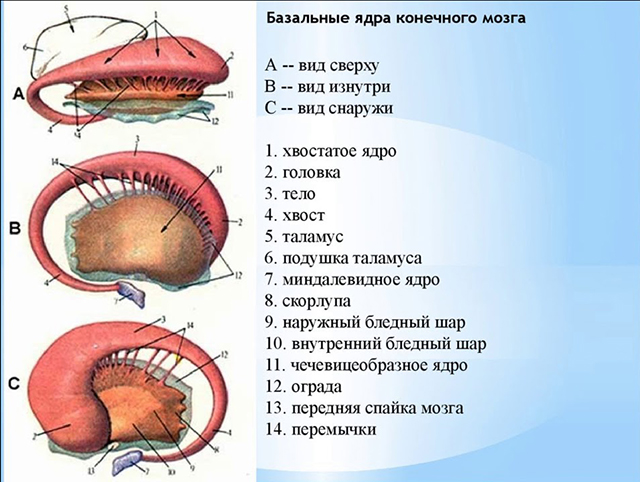
Рис.2.Базальные ядра конечного мозга (полусхематично)
А - вид сверху B -- вид изнутри C -- вид снаружи 1. хвостатое ядро 2. головка 3. тело 4. хвост 5. таламус 6. подушка таламуса 7. миндалевидное ядро 8. скорлупа 9. наружный бледный шар 10. внутренний бледный шар 11. чечевицеобразное ядро 12. ограда 13. передняя спайка мозга 14. перемычки
В противоположность полосатому телу, бледный шар имеет преимущественно крупные нейроны. Кроме того, имеется значительное количество мелких нейронов, выполняющих, по-видимому, функции промежуточных элементов. (Ноздрачева А.Д., 1991)
Ограда содержит полиморфные нейроны разных типов. (Покровский, 1997)
Функции неостриатума
Функции любых образований головного мозга определяется, прежде всего, их связями с неостриатумом. Базальные ганглии образуют многочисленные связи как между структурами входящими в их состав, так и другими отделами мозга. Эти связи представлены в виде параллельных петель, связывающих кору больших полушарий (двигательную, соматосенсорную, лобную) с таламусом. Информация поступает из вышеперечисленных зон коры, проходит через базальные ядра (хвостатое ядро и скорлупу) и черное вещество в двигательные ядра таламуса оттуда снова возвращается в эти же зоны коры - это скелетомоторная петля. Одна из таких петель управляет движениями лица и рта, контролирует такие параметры движения как сила, амплитуда и направление.
Другая петля - глазодвигательная (окуломоторная) специализируется на движении глаза (Агаджанян Н.А., 2001)
Неостриатум имеет также функциональные связи со структурами, лежащими вне этого круга: с черной субстанцией, красным ядром, вестибулярными ядрами, мозжечком, мотонейронами спинного мозга.
Обилие и характер связей неостриатума свидетельствует о его участии в интегративных процессах (аналитикосинтетическая деятельность, обучение, память, рассудок, речь, сознание), в организации и регуляции движений, регуляции работы вегетативных органов.
Некоторые из этих структур, например, черная субстанция, оказывает модулирующее влияние на хвостатое ядро. Взаимодействие черной субстанции с неостриатумом основано на прямых и обратных связях между ними. Стимуляция хвостатого ядра усиливает активность нейронов черного вещества. Стимуляция черного вещества приводит к увеличению, а его разрушение - уменьшает количество дофамина в хвостатом ядре. Дофамин синтезируется в клетках черной субстанции, а затем со скоростью 0,8 мм в час транспортируется к синапсам нейронов хвостатого ядра. В неостриатуме на 1 г нервной ткани накапливается до 10 мг дофамина, что в 6 раз больше, чем в других отделах переднего мозга, например в бледном шаре и в 19 раз больше, чем в мозжечке. Дофамин подавляет фоновую активность большинства нейронов хвостатого ядра, а это позволяет снять тормозящее действие этого ядра на активность бледного шара. Благодаря дофамину появляется растормаживающий механизм взаимодействия между нео- и палеостриатумом. При недостатке дофамина в неостриатуме, что наблюдается при дисфункции черного вещества, нейроны бледного шара растормаживаются, активизируют спинно-стволовые системы, это приводит к двигательным нарушениям в виде ригидности мышцы.
Во взаимодействиях неостриатума и палеостриатума между собой превалируют тормозные влияния. Если раздражать хвостатое ядро, то большая часть нейронов бледного шара тормозится, часть вначале возбуждается - затем тормозится, меньшая часть нейронов возбуждается.
Неостриатум и палеостриатум принимают участие в таких интегративных процессах как условнорефлекторная деятельность, двигательная активность. Это выявляется при их стимуляции, деструкции и при регистрации электрической активности.
Прямое раздражение некоторых зон неостриатума вызывает поворот головы в сторону, противоположную раздражаемому полушарию, животное начинает двигаться по кругу, т.е. возникает так называемая циркуляторная реакция. Раздражение других областей неостриатума вызывает прекращение всех видов активности человека или животного: ориентировочной, эмоциональной, двигательной, пищевой. При этом в коре мозга наблюдается медленно-волновая электрическая активность.
У человека во время нейрохирургической операции, стимуляция хвостатого ядра нарушает речевой контакт с больным: если больной что-то говорил, то он замолкает, а после прекращения раздражения не помнит, что к нему обращались. В случае травм черепа с симптомами раздражения неостриатума у больных отмечается ретро-, антеро- или ретроантероградная амнезия -выпадение памяти на событие, предшествующее травме. Раздражение хвостатого ядра на разных этапах выработки рефлекса приводит к торможению выполнения этого рефлекса.
Раздражение хвостатого ядра может полностью предотвратить восприятие болевых, зрительных, слуховых и других видов стимуляции. Раздражение вентральной области хвостатого ядра снижает, а дорсальной повышает слюноотделение.
Ряд подкорковых структур так же получает тормозное влияние со стороны хвостатого ядра. Так, стимуляция хвостатых ядер вызывала веретенообразную активность в зрительном бугре, бледном шаре, субталамическом теле, черном веществе и др.
Таким образом, специфичным для раздражения хвостатого ядра является торможение активности коры, подкорки, торможение безусловного и условно-рефлекторного поведения.
Хвостатое ядро имеет наряду с тормозящими структурами и возбуждающие. Поскольку возбуждение неостриатума тормозит движения, вызываемые с других пунктов мозга, то оно может тормозить и движения, вызываемые раздражением самого неостриатума. В то же время, если его возбудительные системы стимулируются изолированно, они вызывают то или иное движение. Если считать, что функции хвостатого ядра заключается в обеспечении перехода одного вида движения в другое, т.е прекращение одного движения и обеспечении нового путем создания позы, условий для изолированных движений, то становится понятным существование двух функций хвостатого ядра - тормозной и возбуждающей.
Эффекты выключения неостриатума показали, что функция его ядер связана с регуляцией тонуса мускулатуры. Так, при повреждении этих ядер наблюдались гиперкинезы типа непроизвольных мимических реакций, тремора, торсионного спазма, хореи (подергивания конечностей, туловища, как при нескоординированном танце), двигательной гиперактивности в форме бесцельного перемещения с места на место.
При повреждении неостриатума имеет место расстройства высшей нервной деятельности, затруднение ориентации в пространстве, нарушение памяти, замедление роста организма. После двустороннего повреждения хвостатого ядра условные рефлексы исчезают на длительный срок, выработка новых рефлексов затрудняется, дифференцировка, если и образуется, то отличается непрочностью, отсроченные реакции выработать не удается.
При повреждении хвостатого ядра общее поведение отличается застойностью, инертностью, трудностью переключений с одной формы поведения на другую. При воздействиях на хвостатое ядро имеют места расстройства движения: двустороннее повреждение полосатого тела ведет к безудержному стремлению движения вперед, одностороннее повреждение приводит к манежным движениям.
Несмотря на большое функциональное сходство хвостатого ядра и скорлупы, все же имеет ряд функций, специфичных для последней. Для скорлупы характерно участие в организации пищевого поведения; ряд трофических нарушений кожи, внутренних органов (например, гепатолентикулярная дегенерация) возникает при дефиците функции скорлупы. Раздражения скорлупы приводят к изменениям дыхания, слюноотделения.
Из фактов о том, что стимуляция неостриатума приводит к торможению условного рефлекса, следовало бы ожидать, что разрушение хвостатого ядра вызовет облегчение условнорефлекторной деятельности. Но оказалось, что разрушение хвостатого ядра также приводит к торможению условнорефлекторной деятельности. Видимо, функция хвостатого ядра не является просто тормозной, а заключается в корреляции и интеграции процессов оперативной памяти. Об этом свидетельствует также тот факт, что на нейронах хвостатого ядра конвергирует информация различных сенсорных систем, так как большая часть этих нейронов полисенсорна. Таким образом, неостриатум является подкорковым интегративным и ассоциативным центром.
Функции палеостриатума (бледного шара)
В отличие от неостриатума, стимуляция палеостриатума не вызывает торможения, а провоцирует ориентировочную реакцию, движение конечностей, пищевое поведение (жевание, глотание). Разрушение бледного шара приводит к гипомимии (маскообразное лицо), гиподинамии, эмоциональной тупости. Повреждение бледного шара вызывает у людей тремор головы, конечностей, причем этот тремор исчезает в покое, во время сна и усиливается при движении конечностей, речь становится монотонной. При повреждении бледного шара имеет место миоклония - быстрые подергивания отдельных мышечных групп или отдельных мышц рук, спины, лица. У человека с дисфункцией бледного шара начало движений становится трудным, исчезают вспомогательные и реактивные движения при вставании, нарушаются содружественные помахивания рук при ходьбе.
Функции ограды
Ограда тесно связана с островской корой как прямыми, так и обратными связями. Кроме того, прослеживаются связи ограды к лобной, затылочной, височной коре, показаны обратные связи от коры к ограде. Ограда связана с обонятельной луковицей, с обонятельной корой своей и контралатеральной стороны, а также с оградой другого полушария. Из подкорковых образований ограда связана со скорлупой, хвостатым ядром черным веществом, миндалевидным комплексом, зрительным бугром, бледным шаром.
Реакции нейронов ограды широко представлены на соматические, слуховые, зрительные раздражения, причем эти реакции, в основном, возбудительного характера. Атрофия ограды приводит к полной потере способности больного говорить. Стимуляция ограды вызывает ориентировочную реакцию, поворот головы, жевательные, глотательные, иногда рвотные движения. Эффекты раздражения ограды на условный рефлекс, предъявление стимуляции в разные фазы условного рефлекса тормозит условный рефлекс на счет, мало сказывается при условном рефлексе на звук. Если раздражение производилось одновременно с подачей условного сигнала, то условный рефлекс тормозился. Стимуляция ограды во время еды тормозит поведение пищи. При повреждении ограды левого полушария у человека наблюдается расстройство речи.
Таким образом базальные ганглии головного мозга являются интегративными центрами организации моторики, эмоции, высшей нервной деятельности. Причем, каждая из этих функций может быть усилена или заторможена активацией отдельных образований базальных ядер. (Ткаченко, 1994)
кишка мембранный мозг неостриатум





