Клеточная (плазматическая) мембрана, ее основные функции. Строение и функции биологических мембран. Транспорт веществ через биологические мембраны
Мембраны – это чрезвычайно вязкие и вместе с тем пластичные структуры, окружающие все живые клетки. Функции клеточных мембран:
1.Плазматическая мембрана является барьером, с помощью которого поддерживается различный состав вне- и внутриклеточной среды.
2.Мембраны формируют специализированные компартменты внутри клетки, т.е. многочисленные органеллы – митохондрии, лизосомы, комплекс Гольджи, эндоплазматический ретикулум, ядерные мембраны.
3.В мембранах локализованы ферменты, участвующие в преобразовании энергии в таких процессах, как окислительное фосфорилирование и фотосинтез.
Строение и состав мембран
Основу мембраны составляет двойной липидный слой, в формировании которого участвуют фосфолипиды и гликолипиды. Липидный бислой образован двумя рядами липидов, гидрофобные радикалы которых спрятаны внутрь, а гидрофильные группы обращены наружу и контактируют с водной средой. Белковые молекулы как бы “растворены” в липидном бислое.
Структура липидов мембран
Мембранные липиды − амфифильные молекулы, т.к. в молекуле есть как гидрофильный участок (полярные головки), так и гидрофобный участок, представленный углеводородными радикалами жирных кислот, самопроизвольно формирующие бислой. В мембранах присутствуют липиды трех главных типов – фосфолипиды, гликолипиды и холестерол.
Липидный состав различен. Содержание того или иного липида, по-видимому, определяется разнообразием функций, выполняемых этими липидами в мембранах.
Фосфолипиды. Все фосфолипиды можно разделить на две группы – глицерофосфолипиды и сфингофосфолипиды. Глицерофосфолипиды относят к производным фосфатидной кислоты. Наиболее распространенные глицерофосфолипиды – фосфатидилхолины и фосфатидилэтаноламины. Сфингофосфолипиды построены на основе аминоспирта сфингозина.
Гликолипиды. В гликолипидах гидрофобная часть представлена спиртом церамидом, а гидрофильная – углеводным остатком. В зависимости от длины и строения углеводной части различают цереброзиды и ганглиозиды. Полярные “головки” гликолипидов находятся на наружной поверхности плазматических мембран.
Холестерол (ХС). ХС присутствует во всех мембранах животных клеток. Его молекула состоит из жесткого гидрофобного ядра и гибкой углеводородной цепи. Единственная гидроксильная группа в 3-положении является “полярной головкой”. Для животной клетки среднее молярное отношение ХС/фосфолипиды равно 0,3-0,4, но в плазматической мембране это отношение гораздо выше (0,8-0,9). Наличие ХС в мембранах уменьшает подвижность жирных кислот, снижает латеральную диффузию липидов и поэтому может влиять на функции мембранных белков.
Свойства мембран :
1. Избирательная проницаемость. Замкнутый бислой обеспечивает одно из основных свойств мембраны: он непроницаем для большинства водорастворимых молекул, поскольку они не растворяются в его гидрофобной сердцевине. Способностью легко проникать в клетку обладают газы, такие как кислород, СО 2 и азот вследствие малого размера молекул и слабого взаимодействия с растворителями. Также без труда проникают через бислой молекулы липидной природы, например, стероидные гормоны.
2.Жидкостность. Для мембран характерна жидкостность (текучесть), способность липидов и белков к перемещениям. Возможны два типа перемещений фосфолипидов – это кувырок (в научной литературе называется “флип-флоп”) и латеральная диффузия. В первом случае противостоящие друг другу в бимолекулярном слое молекулы фосфолипидов переворачиваются (или совершают кувырок) навстречу друг другу и меняются местами в мембране, т.е. наружная становится внутренней и наоборот. Такие перескоки связаны с затратой энергии. Чаще наблюдаются повороты вокруг оси (ротация) и латеральная диффузия – перемещение в пределах слоя параллельно поверхности мембраны. Скорость перемещения молекул зависит от микровязкости мембран, которая, в свою очередь определяется относительным содержанием насыщенных и ненасыщенных жирных кислот в составе липидов. Микровязкость меньше, если в составе липидов преобладают ненасыщенные жирные кислоты, и больше при высоком содержании насыщенных жирных кислот.
3.Асимметрия мембран. Поверхности одной и той же мембраны различаются по составу липидов, белков и углеводов (поперечная асимметрия). Например, в наружном слое преобладают фосфатидилхолины, а во внутреннем – фосфатидилэтаноламины и фосфатидилсерины. Углеводные компоненты гликопротеинов и гликолипидов выходят на наружную поверхность, образуя сплошное поурытие, называемое гликокаликсом. На внутренней поверхности углеводы отсутствуют. Белки – рецепторы гормонов располагаются на наружной поверхности плазматической мембраны, а регулируемые ими ферменты – аденилатциклаза, фосфолипаза С – на внутренней и т.д.
Мембранные белки
Мембранные фосфолипиды играют роль растворителя для мембранных белков, создавая микроокружение, в котором последние могут функционировать. На долю белков приходится от 30 до 70% массы мембран. Число разных белков в мембране варьирует от 6-8 в саркоплазматическом ретикулуме до более чем 100 в плазматической мембране. Это ферменты, транспортные белки, структурные белки, антигены, в том числе антигены основной системы гистосовместимости, рецепторы для различных молекул.
По локализации в мембране белки подразделяются на интегральные (частично или полностью погруженные в мембрану) и периферические (расположенные на ее поверхности). Некоторые интегральные белки пронизывают мембрану один раз (гликофорин), другие прошивают мембрану многократно. Например, фоторецептор сетчатки глаза и β 2 -адренорецептор пересекает бислой 7 раз.
Периферические белки и домены интегральных белков, расположенные на наружной поверхности всех мембран, почти всегда гликозилированы. Олигосахаридные остатки защищают белок от протеолиза, а также участвуют в узнавании лигандов или адгезии.
Клеточные мембраны: их структура и функции
Мембраны – это чрезвычайно вязкие и вместе с тем пластичные структуры, окружающие все живые клетки. Функции клеточных мембран:
1.Плазматическая мембрана является барьером, с помощью которого поддерживается различный состав вне- и внутриклеточной среды.
2.Мембраны формируют специализированные компартменты внутри клетки, т.е. многочисленные органеллы – митохондрии, лизосомы, комплекс Гольджи, эндоплазматический ретикулум, ядерные мембраны.
3.В мембранах локализованы ферменты, участвующие в преобразовании энергии в таких процессах, как окислительное фосфорилирование и фотосинтез.
Структура мембран
В 1972 году Сингер и Николсон предложили жидкостно-мозаичную модель мембранной структуры. Согласно этой модели функционирующие мембраны представляют собой двумерный раствор глобулярных интегральных белков, растворенных в жидком фосфолипидном матриксе. Таким образом, основу мембран составляет бимолекулярный липидный слой, с упорядоченным расположением молекул.
При этом гидрофильный слой образован полярной головкой фосфолипидов (фосфатным остатком, с присоединенным к нему холином, этаноламином или серином) а также углеводной частью гликолипидов. А гидрофобный слой – углеводородными радикалами жирных кислот и сфингозина фосфолипидов и гликолипидов.
Свойства мембран:
1. Избирательная проницаемость. Замкнутый бислой обеспечивает одно из основных свойств мембраны: он непроницаем для большинства водорастворимых молекул, поскольку они не растворяются в его гидрофобной сердцевине. Способностью легко проникать в клетку обладают газы, такие как кислород, СО 2 и азот вследствие малого размера молекул и слабого взаимодействия с растворителями. Также без труда проникают через бислой молекулы липидной природы, например, стероидные гормоны.
2.Жидкостность. Двойной липидный слой имеет жидкокристаллическую структуру, поскольку в целом липидная прослойка жидкая, но в ней есть участки затвердевания, похожие на кристаллические структуры. Хотя положение молекул липидов упорядочено, они сохраняют способность к перемещениям. Возможны два типа перемещений фосфолипидов – это кувырок (в научной литературе называется “флип-флоп”) и латеральная диффузия. В первом случае противостоящие друг другу в бимолекулярном слое молекулы фосфолипидов переворачиваются (или совершают кувырок) навстречу друг другу и меняются местами в мембране, т.е. наружная становится внутренней и наоборот. Такие перескоки связаны с затратой энергии и совершаются очень редко. Чаще наблюдаются повороты вокруг оси (ротация) и латеральная диффузия – перемещение в пределах слоя параллельно поверхности мембраны.
3.Асимметрия мембран. Поверхности одной и той же мембраны различаются по составу липидов, белков и углеводов (поперечная асимметрия). Например, в наружном слое преобладают фосфатидилхолины, а во внутреннем – фосфатидилэтаноламины и фосфатидилсерины. Углеводные компоненты гликопротеинов и гликолипидов выходят на наружную поверхность, образуя сплошное поурытие, называемое гликокаликсом. На внутренней поверхности углеводы отсутствуют. Белки – рецепторы гормонов располагаются на наружной поверхности плазматической мембраны, а регулируемые ими ферменты – аденилатциклаза, фосфолипаза С – на внутренней и т.д.
Мембранные белки
Мембранные фосфолипиды играют роль растворителя для мембранных белков, создавая микроокружение, в котором последние могут функционировать. Число разных белков в мембране варьирует от 6-8 в саркоплазматическом ретикулуме до более чем 100 в плазматической мембране. Это ферменты, транспортные белки, структурные белки, антигены, в том числе антигены основной системы гистосовместимости, рецепторы для различных молекул.
По локализации в мембране белки подразделяются на интегральные (частично или полностью погруженные в мембрану) и периферические (расположенные на ее поверхности). Некоторые интегральные белки прошивают мембрану многократно. Например, фоторецептор сетчатки глаза и β 2 -адренорецептор пересекает бислой 7 раз.
Перенос вещества и информации через мембраны
Клеточные мембраны не являются наглухо закрытыми перегородками. Одной из основных функций мембран является регуляция переноса веществ и информации. Трансмембранное перемещение малых молекул осуществляется 1) путем диффузии, пассивной или облегченной и 2) путем активного транспорта. Трансмембранное перемещение крупных молекул осуществляется 1) путем эндоцитоза и 2) путем экзоцитоза. Передача сигнала через мембраны осуществляется с помощью рецепторов, локализованных на наружной поверхности плазматической мембраны. При этом сигнал либо подвергается трансформации (например, глюкагон цАМФ), либо происходит его интернализация, сопряженная с эндоцитозом (например, ЛНП - рецептор ЛНП).
Простая диффузия - это проникновение в клетку веществ по электрохимическому градиенту. При этом никаких энергетических затрат не требуется. Скорость простой диффузии определяется 1) трансмембранным концентрационным градиентом вещества и 2) его растворимостью в гидрофобном слое мембраны.
При облегченной диффузии вещества переносятся через мембрану также по градиенту концентрации, без энергетических затрат, но с помощью специальных мембранных белков-переносчиков. Поэтому облегченная диффузия по ряду параметров отличается от пассивной: 1) для облегченной диффузии характерна высокая избирательность, т.к. белок-переносчик имеет активный центр, комплементарный переносимому веществу; 2) скорость облегченной диффузии способна выходить на плато, т.к. количество молекул-переносчиков ограничено.
Одни транспортные белки просто переносят какое-либо вещество с одной стороны мембраны на другую. Такой простой перенос называется пассивным унипортом. Примером унипорта могут служить ГЛЮТ – транспортеры глюкозы, осуществляющие транспорт глюкозы через клеточные мембраны. Другие белки функционируют как ко-транспортные системы, в которых перенос одного вещества зависит от одновременного или последовательного переноса другого вещества либо в том же направлении – такой перенос называется пассивным симпортом, либо в противоположном направлении – такой перенос называется пассивным антипортом. По механизму пассивного антипорта функционируют транслоказы внутренней мембраны митохондрий, в частности, АДФ/АТФ-транслоказа.
При активном транспорте перенос вещества осуществляется против градиента концентрации и поэтому сопряжен с энергетическими затратами. Если перенос лигандов через мембрану связан с затратой энергии АТФ, то такой перенос называется первично-активным транспортом. Примером могут служить Na + K + -АТФаза и Са 2+ -АТФаза, локализованные в плазматической мембране клеток человека и Н + ,К + -АТФаза слизистой оболочки желудка.
Вторично-активный транспорт. Перенос некоторых веществ против градиента концентрации зависит от одновременного или последовательного переноса Na + (ионов натрия) по градиенту концентрации. При этом, если лиганд переносится в том же направлении, что и Na + , процесс называется активным симпортом. По механизму активного симпорта происходит всасывание глюкозы из просвета кишечника, где ее концентрация низка. Если же лиганд переносится в противоположном ионам натрия направлении, то такой процесс называется активным антипортом. Примером может служить Na + ,Ca 2+ -обменник плазматической мембраны.
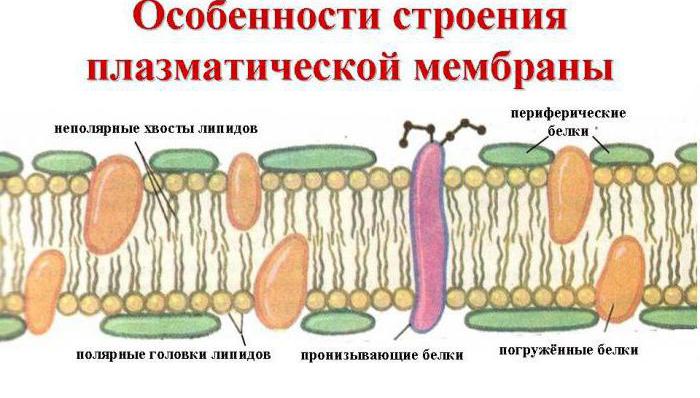
Плазматическая мембрана , или плазмалемма, - наиболее постоянная, основная, универсальная для всех клеток мембрана. Она представляет собой тончайшую (около 10 нм) пленку, покрывающую всю клетку. Плазмалемма состоит из молекул белков и фосфолипидов (рис. 1.6).
Молекулы фосфолипидов расположены в два ряда - гидрофобными концами внутрь, гидрофильными головками к внутренней и внешней водной среде. В отдельных местах бислой (двойной слой) фосфолипидов насквозь пронизан белковыми молекулами (интегральные белки). Внутри таких белковых молекул имеются каналы - поры, через которые проходят водорастворимые вещества. Другие белковые молекулы пронизывают бислой липидов наполовину с одной или с другой стороны (полуинтегральные белки). На поверхности мембран эукариотических клеток имеются периферические белки. Молекулы липидов и белков удерживаются благодаря гидрофильно-гидрофобным взаимодействиям.
Свойства и функции мембран. Все клеточные мембраны представляют собой подвижные текучие структуры, поскольку молекулы липидов и белков не связаны между собой ковалентными связями и способны достаточно быстро перемещаться в плоскости мембраны. Благодаря этому мембраны могут изменять свою конфигурацию, т. е. обладают текучестью.
Мембраны - структуры очень динамичные. Они быстро восстанавливаются после повреждения, а также растягиваются и сжимаются при клеточных движениях.
Мембраны разных типов клеток существенно различаются как по химическому составу, так и по относительному содержанию в них белков, гликопротеинов, липидов, а следовательно, и по характеру имеющихся в них рецепторов. Каждый тип клеток поэтому характеризуется индивидуальностью, которая определяется в основном гликопротеинами. Разветвленные цепи гликопротеинов, выступающие из клеточной мембраны, участвуют в распознава-нии факторов внешней среды, а также во взаимном узнавании родственных клеток. Например, яйцеклетка и сперматозоид узнают друг друга по гликопротеинам клеточной поверхности, которые подходят другкдругу как отдельные элементы цельной структуры. Такое взаимное узнавание - необходимый этап, предшествующий оплодотворению.
Подобное явление наблюдается в процессе дифференциров-ки тканей. В этом случае сходные по строению клетки с помощью распознающих участков плазмалеммы правильно ориентируются относительно друг друга, обеспечивая тем самым их сцепление и образование тканей. С распознаванием связана и регуляция транспорта молекул и ионов через мембрану, а также иммунологический ответ, в котором гликопротеины играют роль антигенов. Сахара, таким образом, могут функционировать как информационные молекулы (подобно белкам и нуклеиновым кислотам). В мембранах содержатся также специфические рецепторы, переносчики электронов, преобразователи энергии, ферментные белки. Белки участвуют в обеспечении транспорта определенных молекул внутрь клетки или из нее, осуществляют структурную связь цитоскелета с клеточными мембранами или же служат в качестве рецепторов для получения и преобразования химических сигналов из окружающей среды.
Важнейшим свойством мембраны является также избирательная проницаемость. Это значит, что молекулы и ионы проходят через нее с различной скоростью, и чем больше размер молекул, тем меньше скорость прохождения их через мембрану. Это свойство определяет плазматическую мембрану как осмотический барьер. Максимальной проникающей способностью обладает вода и растворенные в ней газы; значительно медленнее проходят сквозь мембрану ионы. Диффузия воды через мембрану называется осмосом.
Существует несколько механизмов транспорта веществ через мембрану.
Диффузия -проникновение веществ через мембрану по градиенту концентрации {из области, где их концентрация выше, в область, где их концентрация ниже). Диффузный транспорт веществ (воды, ионов) осуществляется при участии белков мембраны, в которых имеются молекулярные поры, либо при участии липидной фазы (для жирорастворимых веществ).
При облегченной диффузии специальные мембранные белки-переносчики избирательно связываются с тем или иным ионом или молекулой и переносят их через мембрану по градиенту концентрации.
Активный транспорт сопряжен с затратами энергии и служит для переноса веществ против их градиента концентрации. Он осуществляется специальными белками-переносчиками, образующими так называемыеионные насосы. Наиболее изученным является Na - / К - -насос в клетках животных, активно выкачивающих ионы Na + наружу, поглощая при этом ионы К - . Благодаря этому в клетке поддерживается большая концентрация К - и меньшая Na + по сравнению с окружающей средой. На этот процесс затрачивается энергия АТФ.
В результате активного транспорта с помощью мембранного насоса в клетке происходит также регуляция концентрации Mg 2- и Са 2+ .
В процессе активного транспорта ионов в клетку через цито-плазматическую мембрану проникают различные сахара, нукле-отиды, аминокислоты.
Макромолекулы белков, нуклеиновых кислот, полисахаридов, липопротеидные комплексы и др. сквозь клеточные мембраны не проходят, в отличие от ионов и мономеров. Транспорт макромолекул, их комплексов и частиц внутрь клетки происходит совершенно иным путем - посредством эндоцитоза. При эндоци-тозе {эндо... - внутрь) определенный участок плазмалеммы захватывает и как бы обволакивает внеклеточный материал, заключая его в мембранную вакуоль, возникшую вследствие впя-чивания мембраны. В дальнейшем такая вакуоль соединяется с лизосомой, ферменты которой расщепляют макромолекулы до мономеров.
Процесс, обратный эндоцитозу, - экзоцитоз (экзо... - наружу). Благодаря ему клетка выводит внутриклеточные продукты или непереваренные остатки, заключенные в вакуоли или пу-
зырьки. Пузырек подходит к цитоплазматической мембране, сливается с ней, а его содержимое выделяется в окружающую среду. Гак выводятся пищеварительные ферменты, гормоны, гемицел-люлоза и др.
Таким образом, биологические мембраны как основные структурные элементы клетки служат не просто физическими границами, а представляют собой динамичные функциональные поверхности. На мембранах органелл осуществляются многочисленные биохимические процессы, такие как активное поглощение веществ, преобразование энергии, синтез АТФ и др.
Функции биологических мембран следующие:
Отграничивают содержимое клетки от внешней среды и содержимое органелл от цитоплазмы.
Обеспечивают транспорт веществ в клетку и из нее, из цитоплазмы в органеллы и наоборот.
Выполняют роль рецепторов (получение и преобразование сит-налов из окружающей среды, узнавание веществ клеток и т. д.).
Являются катализаторами (обеспечение примембранных химических процессов).
Участвуют в преобразовании энергии.
По функциональным особенностям клеточную мембрану можно разделить на 9 выполняемых ей функций.
Функции клеточной мембраны:
1. Транспортная. Производит транспорт веществ из клетки в клетку;
2. Барьерная. Обладает избирательной проницаемостью, обеспечивает необходимый обмен веществ;
3. Рецепторная. Некоторые белки находящиеся в мембране являются рецепторами;
4. Механическая. Обеспечивает автономность клетки и её механических структур;
5. Матричная. Обеспечивает оптимальное взаимодействие и ориентацию матричных белков;
6. Энергетическая. В мембранах действуют системы переноса энергии при клеточном дыхании в митохондриях;
7. Ферментативная. Мембранные белки иногда являются ферментами. Например мембраны клеток кишечника;
8. Маркировочная. На мембране есть антигены (гликопротеины), которые позволяют опознать клетку;
9. Генерирующая. Осуществляет генерацию и проведение биопотенциалов.
Посмотреть как выглядит клеточная мембрана можно на примере строения животной клетки или растительной клетки .
На рисунке приведено строение клеточной мембраны.
К компонентам клеточной мембраны можно отнести различные белки клеточной мембраны (глобулярный, переферический, поверхностный), а также липиды клеточной мембраны (гликолипид, фосфолипид). Таже в строении клеточной мембраны присутствуют углеводы, холестерол, гликопротеин и белковая альфа спираль.
Состав клеточной мембраны
К основному составу клеточной мембраны относятся:
1. Белки - отвечающие за разнообразные свойства мембраны;
2. Липиды трёх видов (фосфолипиды, гликолипиды и холестерол) отвечающих за жёсткость мембраны.
Белки клеточной мембраны:
1. Глобулярный белок;
2. Поверхностный белок;
3. Переферический белок.
Основное назначение клеточной мембраны
Основное назначение клеточной мембраны:
1. Регулировать обмен между клеткой и средой;
2. Отделять содержимое любой клетки от внешней среды тем самым обеспечивая её целостность;
3. Внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки - органеллы или компартменты, в которых поддерживаются определённые условия среды.
Структура клеточной мембраны
Структура клеточной мембраны представляют собой двумерный раствор глобулярных интегральных белков, растворенных в жидком фосфолипидном матриксе. Данная модель мембранной структуры была предложена двумя учёными Никольсоном и Сингером в 1972 году. Таким образом, основу мембран составляет бимолекулярный липидный слой, с упорядоченным расположением молекул, что вы могли видеть на .
Изучением строения организмов, а также растений животных и человека занимается раздел биологии, называемый цитологией. Ученые установили, что содержимое клетки, которое находится внутри нее, построено довольно сложно. Его окружает так называемый поверхностный аппарат, в состав которого входят наружная клеточная мембрана, надмембранные структуры: гликокаликс и а также микронити, пеликула и микротрубочки, образующие её подмембранный комплекс.
В данной статье мы изучим строение и функции наружной клеточной мембраны, входящей в поверхностный аппарат различных видов клеток.
Какие функции выполняет наружная клеточная мембрана
Как было описано ранее, наружная мембрана является частью поверхностного аппарата каждой клетки, который успешно отделяет ее внутреннее содержимое и защищает клеточные органеллы от неблагоприятных условий внешней среды. Еще одна функция - это обеспечение обмена веществ между клеточным содержимым и тканевой жидкостью, поэтому наружная клеточная мембрана осуществляет транспорт молекул и ионов, поступающих в цитоплазму, а также помогает удалять шлаки и избыток токсичных веществ из клетки.

Строение клеточной мембраны
Мембраны, или плазмалеммы различных типов клеток сильно отличаются между собой. Главным образом, химическим строением, а также относительным содержанием в них липидов, гликопротеидов, белков и, соответственно, характером рецепторов, находящихся в них. Наружная которой определяются прежде всего индивидуальным составом гликопротеидов, берет участие в распознавании раздражителей внешней среды и в реакциях самой клетки на их действия. С белками и гликолипидами клеточных мембран могут взаимодействовать некоторые виды вирусов, вследствие чего они проникают в клетку. Вирусы герпеса и гриппа могут использовать для построения свой защитной оболочки.
А вирусы и бактерии, так называемые бактериофаги, прикрепляются к мембране клетки и в месте контакта растворяют ее с помощью особого фермента. Затем в образовавшееся отверстие проходит молекула вирусной ДНК.
Особенности строения плазмалеммы эукариот
Напомним, что наружная клеточная мембрана выполняет функцию транспорта, то есть переноса веществ в и из нее во внешнюю среду. Для осуществления такого процесса необходимо специальное строение. Действительно, плазмалемма представляет собой постоянную, универсальную для всех систему поверхностного аппарата. Это тоненькая (2-10 Нм), но достаточно плотная многослойная пленка, которая покрывает всю клетку. Её строение было изучено в 1972 году такими учеными, как Д. Сингер и Г. Николсон, ими же создана жидкостно-мозаичная модель клеточной мембраны.
Главные химические соединения, которые её образуют - это упорядоченно расположенные молекулы белков и определенных фосфолипидов, которые вкраплены в жидковатую липидную среду и напоминают мозаику. Таким образом, клеточная мембрана состоит из двух слоев липидов, неполярные гидрофобные «хвосты» которых находятся внутри мембраны, а полярные гидрофильные головки обращены к цитоплазме клетки и к межклеточной жидкости.
Слой липидов пронизывается крупными белковыми молекулами, образующими гидрофильные поры. Именно через них транспортируются водные растворы глюкозы и минеральных солей. Некоторые белковые молекулы находятся как на внешней, так и на внутренней поверхности плазмалеммы. Таким образом, на наружной клеточной мембране в клетках всех организмов, имеющих ядра, находятся молекулы углеводов, связанные ковалентными связями с гликолипидами и гликопротеидами. Содержание углеводов в клеточных мембранах колеблется от 2 до 10%.

Строение плазмалеммы прокариотических организмов
Наружная клеточная мембрана у прокариот выполняет сходные функции с плазмалеммами клеток ядерных организмов, а именно: восприятие и передача информации, поступающей из внешней среды, транспорт ионов и растворов в клетку и из нее, защита цитоплазмы от чужеродных реагентов извне. Она может образовывать мезосомы - структуры, возникающие при впячивании плазмалеммы внутрь клетки. На них могут находиться ферменты, участвующие в метаболических реакциях прокариот, например, в репликации ДНК, синтезе белков.
Мезосомы также содержат окислительно-восстановительные ферменты, а у фотосинтетиков находятся бактериохлорофилл (у бактерий) и фикобилин (у цианобактерий).
Роль наружных мембран в межклеточных контактах
Продолжая отвечать на вопрос, какие функции выполняет наружная клеточная мембрана, остановимся на ее роли в У растительных клеток в стенках наружной клеточной мембраны образуются поры, переходящие в целлюлозный слой. Через них возможен выход цитоплазмы клетки наружу, такие тонкие каналы называют плазмодесмами.

Благодаря им связь между соседними растительными клетками очень прочная. У клеток человека и животных места контактов соседних клеточных мембран называются десмосомами. Они характерны для эндотелиальных и эпителиальных клеток, а также встречаются у кардиомиоцитов.
Вспомогательные образования плазмалеммы
Разобраться, чем отличаются растительные клетки от животных, помогает изучение особенностей строения их плазмалемм, которые зависят от того, какие функции выполняет наружная клеточная мембрана. Над ней у животных клеток находится слой гликокаликс. Он образован молекулами полисахаридов, связанных с белками и липидами наружной клеточной мембраны. Благодаря гликокаликсу между клетками возникает адгезия (слипание), приводящая к образованию тканей, поэтому он принимает участие в сигнальной функции плазмалеммы - распознавании раздражителей внешней среды.
Как осуществляется пассивный транспорт определенных веществ через клеточные мембраны
Как было уже сказано ранее, наружная клеточная мембрана участвует в процессе транспортировки веществ между клеткой и внешней средой. Существует два вида переноса через плазмалемму: пассивный (дифузионный) и активный транспорт. К первому относится диффузия, облегченная диффузия и осмос. Движение веществ по градиенту концентрации зависит, прежде всего, от массы и величины молекул, проходящих через клеточную мембрану. Например, мелкие неполярные молекулы легко растворяются в среднем липидном слое плазмалеммы, продвигаются через нее и оказываются в цитоплазме.

Крупные молекулы органических веществ проникают в цитоплазму с помощью специальных белков-переносчиков. Они имеют видовую специфичность и, соединяясь с частицей или ионом, без затрат энергии пассивно переносят их через мембрану по градиенту концентрации (пассивный транспорт). Этот процесс лежит в основе такого свойства плазмалеммы, как избирательная проницаемость. В процессе энергия молекул АТФ не используется, и клетка сберегает её на другие метаболические реакции.
Активный транспорт химических соединений через плазмалемму
Так как наружная клеточная мембрана обеспечивает перенос молекул и ионов из внешней среды внутрь клетки и обратно, становится возможным вывод продуктов диссимиляции, являющихся токсинами, наружу, то есть в межклеточную жидкость. происходит против градиента концентрации и требует использования энергии в виде молекул АТФ. В нем также участвуют белки-переносчики, называемые АТФ-азами, являющиеся одновременно и ферментами.

Примером такого транспорта служит натрий-калиевый насос (ионы натрия переходят из цитоплазмы во внешнюю среду, а ионы калия закачиваются в цитоплазму). К нему способны эпителиальные клетки кишечника и почек. Разновидностями такого способа переноса служат процессы пиноцитоза и фагоцитоза. Таким образом, изучив, какие функции выполняет наружная клеточная мембрана, можно установить, что к процессам пино- и фагоцитоза способны гетеротрофные протисты, а также клетки высших животных организмов, например, лейкоциты.
Биоэлектрические процессы в клеточных мембранах
Установлено, что существует разность потенциалов между наружной поверхностью плазмалеммы (она заряжена положительно) и пристеночным слоем цитоплазмы, заряженным отрицательно. Ее назвали потенциалом покоя, и она присуща всем живым клеткам. А нервная ткань имеет не только потенциал покоя, но и способна к проведению слабых биотоков, которое называют процессом возбуждения. Наружные мембраны нервных клеток-нейронов, принимая раздражение от рецепторов, начинают менять заряды: ионы натрия массированно поступают внутрь клетки и поверхность плазмалеммы становится электроотрицательной. А пристеночный слой цитоплазмы вследствие избытка катионов получает положительный заряд. Это объясняет, по какой причине происходит перезарядка наружной клеточной мембраны нейрона, что вызывает проведение нервных импульсов, лежащих в основе процесса возбуждения.





