Клеточная мембрана: строение и функции. Строение и функции биологических мембран. Транспорт веществ через биологические мембраны
Краткое описание:
Сазонов В.Ф. 1_1 Строение клеточной мембраны [Электронный ресурс] // Кинезиолог, 2009-2018: [сайт]. Дата обновления: 06.02.2018..__.201_). _Описано строение и функционирование клеточной мембраны (синонимы: плазмалемма, плазмолемма, биомембрана, клеточная оболочка, наружная клеточная оболочка, мембрана клетки, цитоплазматическая мембрана). Эти начальные сведения необходимы как для цитологии, так и для понимания процессов нервной деятельности: нервного возбуждения, торможения, работы синапсов и сенсорных рецепторов.
Клеточная мембрана (плазма лемма или плазмо лемма)
Определение понятия
Клеточная мембрана (синонимы: плазмалемма, плазмолемма, цитоплазматическая мембрана, биомембрана) - это тройная липопротеиновая (т.е. "жиро-белковая") оболочка, отделяющая клетку от окружающей среды и осуществлящая управляемый обмен и связь между клеткой и окружающей её средой.
Главное в этом определении - не то, что мембрана отделяет клетку от среды, а как раз то, что она соединяет клетку с окружающей средой. Мембрана - это активная структура клетки, она постоянно работает.
Биологическая мембрана - это ультратонкая бимолекулярная пленка фосфолипидов, инкрустированная белками и полисахаридами. Эта клеточная структура лежит в основе барьерных, механических и матричных свойств живого организма (Антонов В.Ф., 1996).
Образное представление о мембране
Мне клеточная мембрана представляетсся в виде решетчатого забора с множеством дверей в нём, который окружает некую территорию. Всякая мелкая живность может через этот забор свободно перемещаться туда и обратно. Но более крупные посетители могут входить только через двери, да и то не всякие. У разных посетителей ключи только от своих дверей, и через чужие двери они проходить не могут. Так вот через этот забор постоянно идут потоки посетителей туда и обратно, потому что главная функция мембраны-забора двойная: отделять территорию от окружающего пространства и в то же время соединять её с окружающим пространством. Для этого и существует в заборе множество отверстий и дверей - !
Свойства мембраны
1. Проницаемость.
2. Полупроницаемость (частичная проницаемость).
3. Избирательная (синоним: селективная) проницаемость.
4. Активная проницаемость (синоним: активный транспорт).
5. Управляемая проницаемость.
Как видим, основное свойство мембраны - это её проницаемость по отношению к различным веществам.
6. Фагоцитоц и пиноцитоз.
7. Экзоцитоз.
8. Наличие электрических и химических потенциалов, точнее разности потенциалов между внутренней и наружной сторонами мембраны. Образно можно сказать, что "мембрана превращает клетку в "электрическую батарейку" с помощью управления ионными потоками" . Подробности: .
9. Изменения электрического и химического потенциала.
10. Раздражимость. Специальные молекулярные рецепторы, находящиеся на мембране, могут соединяться с сигнальными (управляющими) веществами, вследствие чего может меняться состояние мембраны и всей клетки. Молекулярные рецепторы запускают биохимические реакции в ответ на соединение с ними лигандов (управляющих веществ). Важно отметить, что сигнальное вещество воздействует на рецептор снаружи, а изменения продолжаются внутри клетки. Получается, что мембрана передала информацию из окружающей среды во внутреннюю среду клетки.
11. Каталитическая ферментативная активность. Ферменты могут быть встроены в мембрану или связаны с её поверхностью (как внутри, так и снаружи клетки), и там они осуществляют свою ферментативную деятельность.
12. Изменение формы поверхности и её площади. Это позволяет мембране образовывать выросты наружу или, наоборот, впячивания внутрь клетки.
13. Способность образовывать контакты с другими клеточными мембранами.
14. Адгезия - способность прилипать к твёрдым поверхностям.
Краткий список свойств мембраны
- Проницаемость.
- Эндоцитоз, экзоцитоз, трансцитоз.
- Потенциалы.
- Раздражимость.
- Ферментная активность.
- Контакты.
- Адгезия.
Функции мембраны
1. Неполная изоляция внутреннего содержимого от внешней среды.
2. Главное в работе клеточной мембраны - это обмен различными веществами между клеткой и межклеточной средой. Этому служит такое свойство мембраны как проницаемость. Кроме того, мембрана регулирует этот обмен за счёт того, что регулирует свою проницаемость.
3. Ещё одна важная функция мембраны - создание разности химических и электрических потенциалов между её внутренней и наружной сторонами. За счёт этого внутри клетка имеет отрицательный электрический потенциал - .
4. Через мембрану осуществляется также информационный обмен между клеткой и окружающей её средой. Специальные молекулярные рецепторы, расположенные на мембране, могут связываться с управляющими веществами (гормонами, медиаторами, модуляторами) и запускать в клетке биохимические реакции, приводящие к различным изменениям в работе клетки или в её структурах.
Видео: Строение мембраны клетки
Видеолекция: Подробно о строении мембраны и транспорте
Строение мембраны
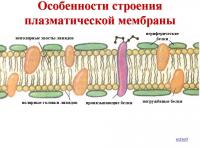
Клеточная мембрана имеет универсальное трёхслойное строение. Её срединный жировой слой является сплошным, а верхний и нижний белковые слои покрывают его в виде мозаики из отдельных белковых участков. Жировой слой является основой, обеспечивающей обособление клетки от окружающей среды, изолирующей её от окружающей среды. Сам по себе он очень плохо пропускает водорастворимые вещества, но легко пропускает жирорастворимые. Поэтому проницаемость мембраны для водорастворимых веществ (например, ионов), приходится обеспечивать специальными белковыми структурами - и .
Ниже представлены микрофотографии реальных клеточных мембран контактирующих клеток, полученные с помощью электронного микроскопа, а также схематический рисунок, показывающий трёхслойность мембраны и мозаичность её белковых слоёв. Для увеличения изображения кликните на него.

Отдельное изображение внутреннего липидного (жирового) слоя клеточной мембраны, пронизанного интегральными встроенными белками. Верхний и нижний белковые слои удалены, чтобы не мешать рассмотрению липидного двойного слоя

Рисунок выше: Неполное схематичное изображение клеточной мембраны (клеточной оболочки), приведённое в Википедии.
Учтите, что наружный и внутренний белковые слои здесь с мембраны сняты, чтобы нам лучше был виден центральный жировой двойной липидный слой. В реальной клеточной мембране сверху и снизу по жировой плёночке (мелкие шарики на рисунке) плавают большие белковые "острова", и мембрана получается более толстой, трёхслойной: белок-жир-белок . Так что она на самом деле похожа на сэндвич из двух белковых "кусков хлеба" с жирным слоем "масла" посередине, т.е. имеет трёхслойное строение, а не двухслойное.
На этом рисунке маленькие голубые и белые шарики соответствуют гидрофильным (смачиваемым) «головкам» липидов, а присоединённые к ним «ниточки» - гидрофобным (несмачиваемым) «хвостам». Из белков показаны только интегральные сквозные мембранные белки (красные глобулы и желтые спирали). Желтые овальные точки внутри мембраны - это молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны - цепочки олигосахаридов , формирующие гликокаликс. Гликокаликс - это как бы углеводный ("сахарный") "пушок" на мембране, образованный торчащими из неё длинными углеводно-белковыми молекулами.
Живая - это маленький «белково-жировой мешочек», заполненный полужидким желеобразным содержимым, которое пронизано плёнками и трубочками.
Стенки этого мешочка образованы двойной жировой (липидной) плёночкой, облепленной изнутри и снаружи белками - клеточной мембраной. Поэтому говорят, что мембрана имеет трёхслойное строение : белки-жиры-белки . Внутри клетки также есть множество подобных жировых мембран, которые делят её внутреннее пространство на отсеки. Такими же мембранами окружены клеточные органеллы: ядро, митохондрии, хлоропласты. Так что мембрана - это универсальная молекулярная структура, свойственная всем клеткам и всем живым организмам.

Слева - уже не реальная, а искусственная модель кусочка биологической мембраны: это мгновенный снимок жирового фосфолипидного бислоя (т.е. двойного слоя) в процессе его молекулярно-динамического моделирования. Показана расчётная ячейка модели - 96 молекул ФХ (ф осфатидилх олина) и 2304 молекулы воды, всего 20544 атомов.
Справа - наглядная модель одиночной молекулы того самого липида, из которых как раз и собирается мембранный липидный бислой. Вверху у него гидрофильная (водолюбивая) головка, а снизу - два гидрофобных (боящихся воды) хвостика. У этого липида есть простое название: 1-стероил-2-докозагексаеноил-Sn-глицеро-3-фосфатидилхолин (18:0/22:6(n-3)cis ФХ), но вам нет нужды его запоминать, если вы только не планируете довести своего преподавателя до обморока глубиной своих познаний.
Можно дать и более точное научное определение клетке:
– это ограниченная активной мембраной, упорядоченная, структурированная неоднородная система биополимеров, участвующих в единой совокупности обменных, энергетических и информационных процессов, и также осуществляющих поддержание и воспроизведение всей системы в целом.
Внутри клетка также пронизана мембранами, а между мембранами находится не вода, а вязкий гель/золь изменяемой плотности. Поэтому взаимодействующие молекулы в клетке не плавают свободно, как в пробирке с водным раствором, а в основном сидят (иммобилизованы) на полимерных структурах цитоскелета или внутриклеточных мембранах. И химические реакции поэтому проходят внутри клетки почти как в твердом теле, а не в жидкости. Наружная мембрана, окружающая клетку, также облеплена ферментами и молекулярными рецепторами, что делает её очень активной частью клетки.
Клеточная мембрана (плазмалемма, плазмолемма) - это активная оболочка, отделяющая клетку от окружающей среды и связывающая её с окружающей средой. © Сазонов В.Ф., 2016.
Из этого определения мембраны следует, что она не просто ограничивает клетку, а активно работает , связывая её с окружающей её средой.
Жир, из которого состоят мембраны, - особенный, поэтому его молекулы принято называть не просто жиром, а «липидами», «фосфолипидами», «сфинголипидами» . Мембранная плёночка является двойной, т. е. она состоит из двух плёночек, слипшихся друг с другом. Поэтому в учебниках пишут, что основа клеточной мембраны состоит из двух липидных слоёв (или из "бислоя ", т.е. двойного слоя). У каждого отдельно взятого липидного слоя одна сторона может смачиваться водой, а другая - не может. Так вот, эти плёночки слипаются друг с другом именно своими несмачивающимися сторонами.
Мембрана бактерий
Оболочка прокариотической клетки грамотрицательных бактерий состоит из нескольких слоёв, показанных на рисунке ниже.
Слои оболочки грамотрицательных бактерий:
1. Внутренняя трёхслойная цитоплазматическая мембрана, которая соприкасается с цитоплазмой.
2. Клеточная стенка, которая состоит из муреина.
3. Наружная трёхслойная цитоплазматическая мембрана, которая имеет такую же систему липидов с белковыми комплексами, как и внутренняя мембрана.
Общение грамотрицательных бактериальных клеток с внешним миром через такую сложную трёхступенчатую структуру не даёт им преимущества в выживании в суровых условиях по сравнению с грамположительным бактериями, имеющими менее мощную оболочку. Они точно так же плохо переносят высокие температуры, повышенную кислотность и перепады давления.




Видеолекция: Плазматическая мембрана. Е.В. Шеваль, к.б.н.
Видеолекция: Мембрана как клеточная граница. А. Иляскин
Важность ионных каналов мембраны
Легко понять, что через мембранную жировую плёнку могут проникать в клетку только жирорастворимые вещества. Это жиры, спирты, газы. Например, в эритроцитх прямо через мембрану легко проходят внутрь и наружу кислород и углекислый газ. А вот вода и водорастворимые вещества (например, ионы) просто так через мембрану не могут пройти внутрь любой клетки. Это значит, что для них нужны специальные отверстия. Но если просто сделать отверстие в жировой плёнке, то оно тут же затянется обратно. Что же делать? Выход в природе был найден: надо сделать специальные белковые транспортные структуры и протянуть их сквозь мембрану. Именно так и получаются каналы для пропускания не растворимых в жире веществ - ионные каналы мембраны клетки.
Итак, для придания своей мембране дополнительных свойства проницаемости для полярных молекул (ионов и воды) клетка синтезирует в цитоплазме специальные белки, которые затем встраиваются в мембрану. Они бывают двух типов: белки-транспортёры (например, транспортные АТФазы) и белки-каналоформеры (образователи каналов). Эти белки встраиваются в двойной жировой слой мембраны и формируют транспортные структуры в виде транспортёров или в виде ионных каналов . Через эти транспортные структуры теперь могут проходить различные водорастворимые вещества, которые по-другому проходить сквозь жировую мембранную плёнку не могут.
Вообще, встроенные в мембрану белки ещё называются интегральными , именно потому что они как бы включаются в состав мембраны и пронизывают её насквозь. Другие белки, не интегральные, образуют как бы острова, «плавающие» по поверхности мембраны: либо по её наружной поверхности, либо по внутренней. Ведь всем известно, что жир является хорошей смазкой и скользить по нему получается легко!
Выводы
1. В целом, мембрана получается трёхслойной:
1) наружный слой из белковых «островов»,
2) жировое двухслойное «море» (липидный бислой), т.е. двойная липидная плёнка,
3) внутренний слой из белковых «островов».
Но есть ещё рыхлый наружный слой - гликокаликс, который образуют торчащие из мембраны гликопротеины. Они являются молекулярными рецепторами, с которыми связываются сигнальные управляющие вещества.
2. В мембрану встроены специальные белковые структуры, обеспечивающие её протицаемость для ионов или других веществ. Не надо забывать, что в некоторых местах жировое море пронизано интегральными белками насквозь. И именно интегральные белки образуют специальные транспортные структуры клеточной мембраны (смотрите раздел 1_2 Транспортные механизмы мембраны). Через них вещества попадают внутрь клетки, а также выводятся из клетки наружу.
3. С любой стороны мембраны (наружной и внутренней), а также внутри мембраны могут располагаться белки-ферменты, которые влияют и на состояние самой мембраны и на жизнь всей клетки.
Так что мембрана клетки - это активная изменчивая структура, которая активно работает в интересах всей клетки и связывает её с окружающим миром, а не просто является "защитной оболочкой". Это - самое важное, что надо знать про клеточную мембрану.
В медицине мембранные белки зачастую используются как “мишени” для лекарственных средств. В качестве таких мишеней выступают рецепторы, ионные каналы, ферменты, транспортные системы. В последнее время кроме мембраны мишенью для лекарственных веществ становятся также гены, спрятанные в клеточном ядре.
Видео: Введение в биофизику клеточной мембраны: Структура мембран 1 (Владимиров Ю.А.)
Видео: История, строение и функции клеточной мембраны: Структура мембран 2 (Владимиров Ю.А.)
© 2010-2018 Сазонов В.Ф., © 2010-2016 kineziolog.bodhy.
Биологические мембраны - общее название функционально активных поверхностных структур, ограничивающих клетки (клеточные, или плазматические мембраны) и внутриклеточные органеллы (мембраны митохондрий, ядер, лизосом, эндоплазматического ретикулума и др.). Они содержат в своем составе липиды, белки,гетерогенные молекулы (гликопротеины,гликолипиды)и в зависимости от выполняемой функции многочисленные минорные компоненты: коферменты, нуклеиновые кислоты, антиоксиданты, каротиноиды, неорганические ионы и т. п.
Согласованное функционирование мембранных систем – рецепторов, ферментов, транспортных механизмов - помогает поддерживать гомеостаз клетки и в то же время быстро реагировать на изменения внешней среды.
К основным функциям биологических мембран можно отнести:
· отделение клетки от окружающей среды и формирование внутриклеточных компартментов (отсеков);
· контроль и регулирование транспорта огромного многообразия веществ через мембраны;
· участие в обеспечении межклеточных взаимодействий, передаче внутрь клетки сигналов;
· преобразование энергии пищевых органических веществ в энергию химических связей молекул АТФ.
Молекулярная организация плазматической (клеточной) мембраны у всех клеток примерно одинакова: она состоит из двух слоев липидных молекул с множеством включенных в нее специфических белков. Одни мембранные белки обладают ферментативной активностью, тогда как другие связывают питательные вещества из окружающей среды и обеспечивают их перенос в клетку через мембраны. Мембранные белки различают по характеру связи с мембранными структурами. Одни белки, называемые внешними или периферическими , непрочно связаны с поверхностью мембраны, другие, называемые внутренними или интегральными , погружены внутрь мембраны. Периферические белки легко экстрагируются, тогда как интегральные белки могут быть выделены только при помощи детергенов или органических растворителей. На рис. 4 представлена структура плазматической мембраны.
Внешние, или плазматические, мембраны многих клеток, а также мембраны внутриклеточных органелл, например, митохондрий, хлоропластов удалось выделить в свободном виде и изучить их молекулярный состав. Во всех мембранах имеются полярные липиды в количестве, составляющем в зависимости от типа мембран от 20 до 80% ее массы, остальное приходится главным образом на долю белков. Так, в плазматических мембранах животных клеток количество белков и липидов, как правило, примерно одинаково; во внутренней митохондриальной мембране содержится около 80% белков и только 20% липидов, а в миелиновых мембранах клеток мозга наоборот, около 80% липидов и только 20% белков.
Рис. 4. Структура плазматической мембраны
Липидная часть мембран представляет собой смесь различного рода полярных липидов. Полярные липиды, к числу которых относятся фосфоглицеролипиды, сфинголипиды, гликолипиды не запасаются в жировых клетках, а встраиваются в клеточные мембраны, причем в строго определенных соотношениях.
Все полярные липиды в мембранах постоянно обновляются в процессе метаболизма, при нормальных условиях в клетке устанавливается динамическое стационарное состояние, при котором скорость синтеза липидов равна скорости их распада.
В мембранах животных клеток присутствуют в основном фосфоглицеролипиды и в меньшей степени сфинголипиды; триацилглицеролы обнаруживаются лишь в следовых количествах. Некоторые мембраны животных клеток, в особенности наружная плазматическая мембрана, содержит значительные количества холестерола и его эфиров (рис.5).

Рис.5. Мембранные липиды
В настоящее время общепринятой моделью строения мембран является жидкостно-мозаичная, предложенная в 1972 году С. Синджером и Дж. Николсоном.
Согласно ей белки можно уподобить айсбергам, плавающим в липидном море. Как уже указывалось выше, существуют 2 типа мембранных белков: интегральные и периферические. Интегральные белки пронизывают мембрану насквозь, они являются амфипатическими молекулами . Периферические белки не пронизывают мембрану и связаны с ней менее прочно. Основной непрерывной частью мембраны, то есть ее матриксом, служит полярный липидный бислой. При обычной для клетки температуре матрикс находится в жидком состоянии, что обеспечивается определенным соотношением между насыщенными и ненасыщенными жирными кислотами в гидрофобных хвостах полярных липидов.
Жидкостно-мозаичная модель предполагает также, что на поверхности расположенных в мембране интегральных белков имеются R-группы аминокислотных остатков (в основном гидрофобные группы, за счет которых белки как бы «растворяются» в центральной гидрофобной части бислоя). В то же время, на поверхности периферических, или внешних белков, имеются в основном гидрофильные R-группы, которые притягиваются к гидрофильным заряженным полярным головкам липидов за счет электростатических сил. Интегральные белки, а к ним относятся ферменты и транспортные белки, обладают активностью только в том случае, если находятся внутри гидрофобной части бислоя, где они приобретают необходимую для проявления активности пространственную конфигурацию (рис.6). Следует еще раз подчеркнуть, что ни между молекулами в бислое, ни между белками и липидами бислоя не образуется ковалентных связей.

Рис.6. Мембранные белки
Мембранные белки могут свободно перемещаться в латериальной плоскости. Периферические белки буквально плавают на поверхности бислойного «моря», а интегральные белки, подобно айсбергам, почти полностью погружены в углеводородный слой.
В большинстве своем мембраны ассиметричны, то есть имеют неравноценные стороны. Эта ассиметричность проявляется в следующем:
· во-первых, в том, что внутренняя и внешняя стороны плазматических мембран бактериальных и животных клеток различаются по составу полярных липидов. Так, например, внутренний липидный слой мембран эритроцитов человека содержит в основном фосфатидилэтаноламин и фосфатидилсерин, а внешний – фосфатидилхолин и сфингомиелин.
· во-вторых, некоторые транспортные системы в мембранах действуют только в одном направлении. Например, в мембранах эритроцитов имеется транспортная система («насос»), перекачивающая ионы Nа + из клетки в окружающую среду, а ионы К + - внутрь клетки за счет энергии, освобождающейся при гидролизе АТФ.
· в-третьих, на внешней поверхности плазматических мембран содержится очень большое число олигосахаридных группировок, представляющих собой головки гликолипидов и олигосахаридные боковые цепи гликопротеинов, тогда как на внутренней поверхности плазматической мембраны олигосахаридных группировок практически нет.
Ассиметричность биологических мембрам сохраняется за счет того, что перенос индивидуальных молекул фосфолипидов с одной стороны липидного бислоя на другую очень затруднен по энергетическим соображениям. Полярная молекула липида способна свободно перемещаться на своей стороне бислоя, но ограничена в возможности перескочить на другую сторону.
Подвижность липидов зависит от относительного содержания и типа присутствующих ненасыщенных жирных кислот. Углеводородная природа жирнокислотных цепей сообщает мембране свойства текучести, подвижности. В присутствии цис-ненасыщенных жирных кислот силы сцепления между цепями слабее, чем в случае одних насыщенных жирных кислот, и липиды сохраняют высокую подвижность и при низкой температуре.
На внешней стороне мембран имеются специфические распознающие участки, функция которых состоит в распознавании определенных молекулярных сигналов. Например, именно посредством мембраны некоторые бактерии воспринимают незначительные изменения концентрации питательного вещества, что стимулирует их движение к источнику пищи; это явление носит название хемотаксиса .
Мембраны различных клеток и внутриклеточных органелл обладают определенной специфичностью, обусловленной их строением, химическим составом и функциями. Выделяют следующие основные группы мембран у эукариотических организмов:
· плазматическая мембрана (наружная клеточная мембрана, плазмалемма),
· ядерная мембрана,
· эндоплазматический ретикулум,
· мембраны аппарата Гольджи, митохондрий, хлорпластов, миелиновых оболочек,
· возбудимые мембраны.
У прокариотических организмов помимо плазматической мембраны существуют внутрицитоплазматические мембранные образования, у гетеротрофных прокариот они называются мезосомами. Последние образуются впячиванием внуть наружной клеточной мембраны и в некоторых случаях сохраняют с ней связь.
Мембрана эритроцитов состоит из белков (50%), липидов (40%) и углеводов (10%). Основная часть углеводов (93%) связана с белками, остальная – с липидами. В мембране липиды расположены асимметрично в отличие от симметричного расположения в мицеллах. Например, кефалин находится преимущественно во внутреннем слое липидов. Такая асимметрия поддерживается, по-видимому, за счет поперечного перемещения фосфолипидов в мембране, осуществляемого с помощью мембранных белков и за счет энергии метаболизма. Во внутреннем слое эритроцитарной мембраны находятся в основном сфингомиелин, фосфатидилэтаноламин, фосфатидилсерин, в наружном слое – фосфатидилхолин. Мембрана эритроцитов содержит интегральный гликопротеин гликофорин , состоящий из 131 аминокислотного остатка и пронизывающий мембрану, и так называемый белок полосы 3, состоящий из 900 аминокислотных остатков. Углеводные компоненты гликофорина выполняют рецепторную функцию для вирусов гриппа, фитогемагглютининов, ряда гормонов. В эритроцитарной мембране обнаружен и другой интегральный белок, содержащий мало углеводов и пронизывающий мембрану. Его называют туннельным белком (компонент а), так как предполагают, что он образует канал для анионов. Периферическим белком, связанным с внутренней стороной эритроцитарной мембраны, является спектрин.
Миелиновые мембраны , окружающие аксоны нейронов, многослойны, в них присутствует большое количество липидов (около 80%, половина из них – фосфолипидов). Белки этих мембран важны для фиксации лежащих друг над другом мембранных солев.
Мембраны хлоропластов . Хлоропласты покрыты двухслойной мембраной. Наружная мембрана имеет некоторое сходство с таковой у митохондрий. Помимо этой поверхностной мембраны в хлоропластах имеется внутренняя мембранная система – ламеллы . Ламеллы образуют или уплощенные пузырьки – тилакоиды, которые, располагаясь друг над другом, собираются в пачки (граны) или формируют мембранную систему стромы (ламеллы стромы). Ламеллы гран и стромы наружной стороне мембраны тилакоидов сосредоточены гидрофильные группировки, галакто- и сульфолипидов. Фитольная часть молекулы хлорофилла погружена в глобулу и находится в контакте в гидрофобными группами белков и липидов. Порфириновые ядра хлорофилла в основном локализованы между соприкасающимися мембранами тилакоидов гран.
Внутренняя (цитоплазматическая) мембрана бактерий по структуре сходна с внутренними мембранами хлоропластов и митохондрий. В ней локализованы ферменты дыхательной цепи, активного транспорта; ферменты, участвующие в образовании компонентов мембраны. Преобладающим компонентом бактериальных мембран являются белки: соотношение белок/липид (по массе) равно 3:1. Наружная мембрана грамотрицательных бактерий по сравнению с цитоплазматической содержит меньшее количество различных фосфолипидов и белков. Обе мембраны различаются по липидному составу. Во внешней мембране находятся белки, образующие поры для проникновения многих низкомолекулярных веществ. Характерным компонентом наружной мембраны является также специфический липополисахарид. Ряд белков наружной мембраны служит рецепторами для фагов.
Мембрана вирусов. Среди вирусов мембранные структуры характерны для содержащих нуклеокапсид, который состоит из белка и нуклеиновой кислоты. Это «ядро» вирусов окружено мембраной (оболочка). Она также состоит из двойного слоя липидов с включенными в него гликопротеинами, расположенными в основном на поверхности мембраны. У ряда вирусов (микровирусы) в мембраны входит 70-80% всех белков, остальные белки содержатся в нуклеокапсиде.
Таким образом, мембраны клеток представляют собой очень сложные структуры; составляющие их молекулярные комплексы образуют упорядоченную двумерную мозаику, что придает поверхности мембран биологическую специфичность.
Клеточные мембраны: их структура и функции
Мембраны – это чрезвычайно вязкие и вместе с тем пластичные структуры, окружающие все живые клетки. Функции клеточных мембран:
1.Плазматическая мембрана является барьером, с помощью которого поддерживается различный состав вне- и внутриклеточной среды.
2.Мембраны формируют специализированные компартменты внутри клетки, т.е. многочисленные органеллы – митохондрии, лизосомы, комплекс Гольджи, эндоплазматический ретикулум, ядерные мембраны.
3.В мембранах локализованы ферменты, участвующие в преобразовании энергии в таких процессах, как окислительное фосфорилирование и фотосинтез.
Структура мембран
В 1972 году Сингер и Николсон предложили жидкостно-мозаичную модель мембранной структуры. Согласно этой модели функционирующие мембраны представляют собой двумерный раствор глобулярных интегральных белков, растворенных в жидком фосфолипидном матриксе. Таким образом, основу мембран составляет бимолекулярный липидный слой, с упорядоченным расположением молекул.
При этом гидрофильный слой образован полярной головкой фосфолипидов (фосфатным остатком, с присоединенным к нему холином, этаноламином или серином) а также углеводной частью гликолипидов. А гидрофобный слой – углеводородными радикалами жирных кислот и сфингозина фосфолипидов и гликолипидов.
Свойства мембран:
1. Избирательная проницаемость. Замкнутый бислой обеспечивает одно из основных свойств мембраны: он непроницаем для большинства водорастворимых молекул, поскольку они не растворяются в его гидрофобной сердцевине. Способностью легко проникать в клетку обладают газы, такие как кислород, СО 2 и азот вследствие малого размера молекул и слабого взаимодействия с растворителями. Также без труда проникают через бислой молекулы липидной природы, например, стероидные гормоны.
2.Жидкостность. Двойной липидный слой имеет жидкокристаллическую структуру, поскольку в целом липидная прослойка жидкая, но в ней есть участки затвердевания, похожие на кристаллические структуры. Хотя положение молекул липидов упорядочено, они сохраняют способность к перемещениям. Возможны два типа перемещений фосфолипидов – это кувырок (в научной литературе называется “флип-флоп”) и латеральная диффузия. В первом случае противостоящие друг другу в бимолекулярном слое молекулы фосфолипидов переворачиваются (или совершают кувырок) навстречу друг другу и меняются местами в мембране, т.е. наружная становится внутренней и наоборот. Такие перескоки связаны с затратой энергии и совершаются очень редко. Чаще наблюдаются повороты вокруг оси (ротация) и латеральная диффузия – перемещение в пределах слоя параллельно поверхности мембраны.
3.Асимметрия мембран. Поверхности одной и той же мембраны различаются по составу липидов, белков и углеводов (поперечная асимметрия). Например, в наружном слое преобладают фосфатидилхолины, а во внутреннем – фосфатидилэтаноламины и фосфатидилсерины. Углеводные компоненты гликопротеинов и гликолипидов выходят на наружную поверхность, образуя сплошное поурытие, называемое гликокаликсом. На внутренней поверхности углеводы отсутствуют. Белки – рецепторы гормонов располагаются на наружной поверхности плазматической мембраны, а регулируемые ими ферменты – аденилатциклаза, фосфолипаза С – на внутренней и т.д.
Мембранные белки
Мембранные фосфолипиды играют роль растворителя для мембранных белков, создавая микроокружение, в котором последние могут функционировать. Число разных белков в мембране варьирует от 6-8 в саркоплазматическом ретикулуме до более чем 100 в плазматической мембране. Это ферменты, транспортные белки, структурные белки, антигены, в том числе антигены основной системы гистосовместимости, рецепторы для различных молекул.
По локализации в мембране белки подразделяются на интегральные (частично или полностью погруженные в мембрану) и периферические (расположенные на ее поверхности). Некоторые интегральные белки прошивают мембрану многократно. Например, фоторецептор сетчатки глаза и β 2 -адренорецептор пересекает бислой 7 раз.
Перенос вещества и информации через мембраны
Клеточные мембраны не являются наглухо закрытыми перегородками. Одной из основных функций мембран является регуляция переноса веществ и информации. Трансмембранное перемещение малых молекул осуществляется 1) путем диффузии, пассивной или облегченной и 2) путем активного транспорта. Трансмембранное перемещение крупных молекул осуществляется 1) путем эндоцитоза и 2) путем экзоцитоза. Передача сигнала через мембраны осуществляется с помощью рецепторов, локализованных на наружной поверхности плазматической мембраны. При этом сигнал либо подвергается трансформации (например, глюкагон цАМФ), либо происходит его интернализация, сопряженная с эндоцитозом (например, ЛНП - рецептор ЛНП).
Простая диффузия - это проникновение в клетку веществ по электрохимическому градиенту. При этом никаких энергетических затрат не требуется. Скорость простой диффузии определяется 1) трансмембранным концентрационным градиентом вещества и 2) его растворимостью в гидрофобном слое мембраны.
При облегченной диффузии вещества переносятся через мембрану также по градиенту концентрации, без энергетических затрат, но с помощью специальных мембранных белков-переносчиков. Поэтому облегченная диффузия по ряду параметров отличается от пассивной: 1) для облегченной диффузии характерна высокая избирательность, т.к. белок-переносчик имеет активный центр, комплементарный переносимому веществу; 2) скорость облегченной диффузии способна выходить на плато, т.к. количество молекул-переносчиков ограничено.
Одни транспортные белки просто переносят какое-либо вещество с одной стороны мембраны на другую. Такой простой перенос называется пассивным унипортом. Примером унипорта могут служить ГЛЮТ – транспортеры глюкозы, осуществляющие транспорт глюкозы через клеточные мембраны. Другие белки функционируют как ко-транспортные системы, в которых перенос одного вещества зависит от одновременного или последовательного переноса другого вещества либо в том же направлении – такой перенос называется пассивным симпортом, либо в противоположном направлении – такой перенос называется пассивным антипортом. По механизму пассивного антипорта функционируют транслоказы внутренней мембраны митохондрий, в частности, АДФ/АТФ-транслоказа.
При активном транспорте перенос вещества осуществляется против градиента концентрации и поэтому сопряжен с энергетическими затратами. Если перенос лигандов через мембрану связан с затратой энергии АТФ, то такой перенос называется первично-активным транспортом. Примером могут служить Na + K + -АТФаза и Са 2+ -АТФаза, локализованные в плазматической мембране клеток человека и Н + ,К + -АТФаза слизистой оболочки желудка.
Вторично-активный транспорт. Перенос некоторых веществ против градиента концентрации зависит от одновременного или последовательного переноса Na + (ионов натрия) по градиенту концентрации. При этом, если лиганд переносится в том же направлении, что и Na + , процесс называется активным симпортом. По механизму активного симпорта происходит всасывание глюкозы из просвета кишечника, где ее концентрация низка. Если же лиганд переносится в противоположном ионам натрия направлении, то такой процесс называется активным антипортом. Примером может служить Na + ,Ca 2+ -обменник плазматической мембраны.
Изучением строения организмов, а также растений животных и человека занимается раздел биологии, называемый цитологией. Ученые установили, что содержимое клетки, которое находится внутри нее, построено довольно сложно. Его окружает так называемый поверхностный аппарат, в состав которого входят наружная клеточная мембрана, надмембранные структуры: гликокаликс и а также микронити, пеликула и микротрубочки, образующие её подмембранный комплекс.
В данной статье мы изучим строение и функции наружной клеточной мембраны, входящей в поверхностный аппарат различных видов клеток.
Какие функции выполняет наружная клеточная мембрана
Как было описано ранее, наружная мембрана является частью поверхностного аппарата каждой клетки, который успешно отделяет ее внутреннее содержимое и защищает клеточные органеллы от неблагоприятных условий внешней среды. Еще одна функция - это обеспечение обмена веществ между клеточным содержимым и тканевой жидкостью, поэтому наружная клеточная мембрана осуществляет транспорт молекул и ионов, поступающих в цитоплазму, а также помогает удалять шлаки и избыток токсичных веществ из клетки.

Строение клеточной мембраны
Мембраны, или плазмалеммы различных типов клеток сильно отличаются между собой. Главным образом, химическим строением, а также относительным содержанием в них липидов, гликопротеидов, белков и, соответственно, характером рецепторов, находящихся в них. Наружная которой определяются прежде всего индивидуальным составом гликопротеидов, берет участие в распознавании раздражителей внешней среды и в реакциях самой клетки на их действия. С белками и гликолипидами клеточных мембран могут взаимодействовать некоторые виды вирусов, вследствие чего они проникают в клетку. Вирусы герпеса и гриппа могут использовать для построения свой защитной оболочки.

А вирусы и бактерии, так называемые бактериофаги, прикрепляются к мембране клетки и в месте контакта растворяют ее с помощью особого фермента. Затем в образовавшееся отверстие проходит молекула вирусной ДНК.
Особенности строения плазмалеммы эукариот
Напомним, что наружная клеточная мембрана выполняет функцию транспорта, то есть переноса веществ в и из нее во внешнюю среду. Для осуществления такого процесса необходимо специальное строение. Действительно, плазмалемма представляет собой постоянную, универсальную для всех систему поверхностного аппарата. Это тоненькая (2-10 Нм), но достаточно плотная многослойная пленка, которая покрывает всю клетку. Её строение было изучено в 1972 году такими учеными, как Д. Сингер и Г. Николсон, ими же создана жидкостно-мозаичная модель клеточной мембраны.
Главные химические соединения, которые её образуют - это упорядоченно расположенные молекулы белков и определенных фосфолипидов, которые вкраплены в жидковатую липидную среду и напоминают мозаику. Таким образом, клеточная мембрана состоит из двух слоев липидов, неполярные гидрофобные «хвосты» которых находятся внутри мембраны, а полярные гидрофильные головки обращены к цитоплазме клетки и к межклеточной жидкости.
Слой липидов пронизывается крупными белковыми молекулами, образующими гидрофильные поры. Именно через них транспортируются водные растворы глюкозы и минеральных солей. Некоторые белковые молекулы находятся как на внешней, так и на внутренней поверхности плазмалеммы. Таким образом, на наружной клеточной мембране в клетках всех организмов, имеющих ядра, находятся молекулы углеводов, связанные ковалентными связями с гликолипидами и гликопротеидами. Содержание углеводов в клеточных мембранах колеблется от 2 до 10%.

Строение плазмалеммы прокариотических организмов
Наружная клеточная мембрана у прокариот выполняет сходные функции с плазмалеммами клеток ядерных организмов, а именно: восприятие и передача информации, поступающей из внешней среды, транспорт ионов и растворов в клетку и из нее, защита цитоплазмы от чужеродных реагентов извне. Она может образовывать мезосомы - структуры, возникающие при впячивании плазмалеммы внутрь клетки. На них могут находиться ферменты, участвующие в метаболических реакциях прокариот, например, в репликации ДНК, синтезе белков.
Мезосомы также содержат окислительно-восстановительные ферменты, а у фотосинтетиков находятся бактериохлорофилл (у бактерий) и фикобилин (у цианобактерий).
Роль наружных мембран в межклеточных контактах
Продолжая отвечать на вопрос, какие функции выполняет наружная клеточная мембрана, остановимся на ее роли в У растительных клеток в стенках наружной клеточной мембраны образуются поры, переходящие в целлюлозный слой. Через них возможен выход цитоплазмы клетки наружу, такие тонкие каналы называют плазмодесмами.

Благодаря им связь между соседними растительными клетками очень прочная. У клеток человека и животных места контактов соседних клеточных мембран называются десмосомами. Они характерны для эндотелиальных и эпителиальных клеток, а также встречаются у кардиомиоцитов.
Вспомогательные образования плазмалеммы
Разобраться, чем отличаются растительные клетки от животных, помогает изучение особенностей строения их плазмалемм, которые зависят от того, какие функции выполняет наружная клеточная мембрана. Над ней у животных клеток находится слой гликокаликс. Он образован молекулами полисахаридов, связанных с белками и липидами наружной клеточной мембраны. Благодаря гликокаликсу между клетками возникает адгезия (слипание), приводящая к образованию тканей, поэтому он принимает участие в сигнальной функции плазмалеммы - распознавании раздражителей внешней среды.
Как осуществляется пассивный транспорт определенных веществ через клеточные мембраны
Как было уже сказано ранее, наружная клеточная мембрана участвует в процессе транспортировки веществ между клеткой и внешней средой. Существует два вида переноса через плазмалемму: пассивный (дифузионный) и активный транспорт. К первому относится диффузия, облегченная диффузия и осмос. Движение веществ по градиенту концентрации зависит, прежде всего, от массы и величины молекул, проходящих через клеточную мембрану. Например, мелкие неполярные молекулы легко растворяются в среднем липидном слое плазмалеммы, продвигаются через нее и оказываются в цитоплазме.

Крупные молекулы органических веществ проникают в цитоплазму с помощью специальных белков-переносчиков. Они имеют видовую специфичность и, соединяясь с частицей или ионом, без затрат энергии пассивно переносят их через мембрану по градиенту концентрации (пассивный транспорт). Этот процесс лежит в основе такого свойства плазмалеммы, как избирательная проницаемость. В процессе энергия молекул АТФ не используется, и клетка сберегает её на другие метаболические реакции.
Активный транспорт химических соединений через плазмалемму
Так как наружная клеточная мембрана обеспечивает перенос молекул и ионов из внешней среды внутрь клетки и обратно, становится возможным вывод продуктов диссимиляции, являющихся токсинами, наружу, то есть в межклеточную жидкость. происходит против градиента концентрации и требует использования энергии в виде молекул АТФ. В нем также участвуют белки-переносчики, называемые АТФ-азами, являющиеся одновременно и ферментами.

Примером такого транспорта служит натрий-калиевый насос (ионы натрия переходят из цитоплазмы во внешнюю среду, а ионы калия закачиваются в цитоплазму). К нему способны эпителиальные клетки кишечника и почек. Разновидностями такого способа переноса служат процессы пиноцитоза и фагоцитоза. Таким образом, изучив, какие функции выполняет наружная клеточная мембрана, можно установить, что к процессам пино- и фагоцитоза способны гетеротрофные протисты, а также клетки высших животных организмов, например, лейкоциты.
Биоэлектрические процессы в клеточных мембранах
Установлено, что существует разность потенциалов между наружной поверхностью плазмалеммы (она заряжена положительно) и пристеночным слоем цитоплазмы, заряженным отрицательно. Ее назвали потенциалом покоя, и она присуща всем живым клеткам. А нервная ткань имеет не только потенциал покоя, но и способна к проведению слабых биотоков, которое называют процессом возбуждения. Наружные мембраны нервных клеток-нейронов, принимая раздражение от рецепторов, начинают менять заряды: ионы натрия массированно поступают внутрь клетки и поверхность плазмалеммы становится электроотрицательной. А пристеночный слой цитоплазмы вследствие избытка катионов получает положительный заряд. Это объясняет, по какой причине происходит перезарядка наружной клеточной мембраны нейрона, что вызывает проведение нервных импульсов, лежащих в основе процесса возбуждения.
9.5.1. Одна из главных функций мембран - участие в переносе веществ. Этот процесс обеспечивается при помощи трёх основных механизмов: простой диффузией, облегчённой диффузией и активным транспортом (рисунок 9.10). Запомните важнейшие особенности этих механизмов и примеры транспортируемых веществ в каждом случае.
Рисунок 9.10. Механизмы транспорта молекул через мембрану
Простая диффузия - перенос веществ через мембрану без участия специальных механизмов. Транспорт происходит по градиенту концентрации без затраты энергии. Путём простой диффузии транспортируются малые биомолекулы - Н2 О, СО2 , О2 , мочевина, гидрофобные низкомолекулярные вещества. Скорость простой диффузии пропорциональна градиенту концентрации.
Облегчённая диффузия - перенос веществ через мембрану при помощи белковых каналов или специальных белков-переносчиков. Осуществляется по градиенту концентрации без затраты энергии. Транспортируются моносахариды, аминокислоты, нуклеотиды, глицерол, некоторые ионы. Характерна кинетика насыщения - при определённой (насыщающей) концентрации переносимого вещества в переносе принимают участие все молекулы переносчика и скорость транспорта достигает предельной величины.
Активный транспорт - также требует участия специальных белков-переносчиков, но перенос происходит против градиента концентрации и поэтому требует затраты энергии. При помощи этого механизма через клеточную мембрану транспортируются ионы Na+ , K+ , Ca2+ , Mg2+ , через митохондриальную - протоны. Для активного транспорта веществ характерна кинетика насыщения.
9.5.2. Примером транспортной системы, осуществляющей активный транспорт ионов, является Na+ ,K+ -аденозинтрифосфатаза (Na+ ,K+ -АТФаза или Na+ ,K+ -насос). Этот белок находится в толще плазматической мембраны и способен катализировать реакцию гидролиза АТФ. Энергия, выделяемая при гидролизе 1 молекулы АТФ, используется для переноса 3 ионов Na+ из клетки во внеклеточное пространство и 2 ионов К+ в обратном направлении (рисунок 9.11). В результате действия Na+ ,K+ -АТФазы создаётся разность концентраций между цитозолем клетки и внеклеточной жидкостью. Поскольку перенос ионов неэквивалентен, то возникает разность электрических потенциалов. Таким образом, возникает электрохимический потенциал, который складывается из энергии разности электрических потенциалов Δφ и энергии разности концентраций веществ ΔС по обе стороны мембраны.

Рисунок 9.11. Схема Na+ , K+ -насоса.
9.5.3. Перенос через мембраны частиц и высокомолекулярных соединений
Наряду с транспортом органических веществ и ионов, осуществляемым переносчиками, в клетке существует совершенно особый механизм, предназначенный для поглощения клеткой и выведения из неё высокомолекулярных соединений при помощи изменения формы биомембраны. Такой механизм называют везикулярным транспортом .

Рисунок 9.12. Типы везикулярного транспорта: 1 - эндоцитоз; 2 - экзоцитоз.
При переносе макромолекул происходит последовательное образование и слияние окружённых мембраной пузырьков (везикул). По направлению транспорта и характеру переносимых веществ различают следующие типы везикулярного транспорта:
Эндоцитоз (рисунок 9.12, 1) — перенос веществ в клетку. В зависимости от размера образующихся везикул различают:
а) пиноцитоз — поглощение жидкости и растворённых макромолекул (белков, полисахаридов, нуклеиновых кислот) с помощью небольших пузырьков (150 нм в диаметре);
б) фагоцитоз — поглощение крупных частиц, таких, как микроорганизмы или обломки клеток. В этом случае образуются крупные пузырьки, называемые фагосомами диаметром более 250 нм.
Пиноцитоз характерен для большинства эукариотических клеток, в то время как крупные частицы поглощаются специализированными клетками - лейкоцитами и макрофагами. На первой стадии эндоцитоза вещества или частицы адсорбируются на поверхности мембраны, этот процесс происходит без затраты энергии. На следующей стадии мембрана с адсорбированным веществом углубляется в цитоплазму; образовавшиеся локальные впячивания плазматической мембраны отшнуровываются от поверхности клетки, образуя пузырьки, которые затем мигрируют внутрь клетки. Этот процесс связан системой микрофиламентов и является энергозависимым. Поступившие в клетку пузырьки и фагосомы могут сливаться с лизосомами. Содержащиеся в лизосомах ферменты расщепляют вещества, содержащиеся в пузырьках и фагосомах до низкомолекулярных продуктов (аминокислот, моносахаридов, нуклеотидов), которые транспортируются в цитозоль, где они могут быть использованы клеткой.
Экзоцитоз (рисунок 9.12, 2) — перенос частиц и крупных соединений из клетки. Этот процесс, как и эндоцитоз, протекает с поглощением энергии. Основными разновидностями экзоцитоза являются:
а) секреция - выведение из клетки водорастворимых соединений, которые используются или воздействуют на другие клетки организма. Может осуществляться как неспециализированными клетками, так и клетками эндокринных желёз, слизистой желудочно-кишечного тракта, приспособленными для секреции производимых ими веществ (гормонов, нейромедиаторов, проферментов) в зависимости от определённых потребностей организма.
Секретируемые белки синтезируются на рибосомах, связанных с мембранами шероховатого эндоплазматического ретикулума. Затем эти белки транспортируются к аппарату Гольджи, где они модифицируются, концентрируются, сортируются, и затем упаковываются в пузырьки, которые отщепляются в цитозоль и в дальнейшем сливаются с плазматической мембраной, так что содержимое пузырьков оказывается вне клетки.
В отличие от макромолекул, секретируемые частицы малых размеров, например, протоны, транспортируются из клетки при помощи механизмов облегчённой диффузии и активного транспорта.
б) экскреция - удаление из клетки веществ, которые не могут быть использованы (например, удаление в ходе эритропоэза из ретикулоцитов сетчатой субстанции, представляющей собой агрегированные остатки органелл). Механизм экскреции, по-видимому, состоит в том, что вначале выделяемые частицы оказываются в цитоплазматическом пузырьке, который затем сливается с плазматической мембраной.





